molekula DNK. Zgradba molekule DNA. Dedne informacije: shranjevanje in prenašanje. Genetska koda. Veriga DNK Kjer se nahajajo molekule DNK
Nukleinske kisline so visokomolekularne snovi, sestavljene iz mononukleotidov, ki so med seboj povezani v polimerno verigo s 3", 5" fosfodiestrskimi vezmi in so na določen način pakirani v celicah.
Nukleinske kisline so biopolimeri dveh vrst: ribonukleinska kislina (RNA) in deoksiribonukleinska kislina (DNA). Vsak biopolimer je sestavljen iz nukleotidov, ki se razlikujejo po ostanku ogljikovih hidratov (riboza, deoksiriboza) in eni od dušikovih baz (uracil, timin). Po teh razlikah so nukleinske kisline dobile svoje ime.
Zgradba deoksiribonukleinske kisline
Nukleinske kisline imajo primarno, sekundarno in terciarno strukturo.
Primarna struktura DNK
Primarna struktura DNK je linearna polinukleotidna veriga, v kateri so mononukleotidi povezani s 3", 5" fosfodiestrskimi vezmi. Izhodiščni material za sestavljanje verige nukleinska kislina v celici je nukleozid 5"-trifosfat, ki je zaradi odstranitve β in γ ostankov fosforne kisline sposoben pritrditi 3" ogljikov atom drugega nukleozida. Tako je 3" atom ogljika ene deoksiriboze kovalentno povezan s 5" atomom ogljika druge deoksiriboze preko enega samega ostanka fosforne kisline in tvori linearno polinukleotidno verigo nukleinske kisline. Od tod tudi ime: 3", 5" fosfodiesterske vezi. Dušikove baze ne sodelujejo pri povezovanju nukleotidov ene verige (slika 1.).
Takšna povezava med ostankom molekule fosforne kisline enega nukleotida in ogljikovim hidratom drugega povzroči nastanek pentozo-fosfatnega skeleta polinukleotidne molekule, na katerega so ena za drugo ob strani pritrjene dušikove baze. Njihovo zaporedje razporeditve v verigah molekul nukleinskih kislin je strogo specifično za celice različnih organizmov, tj. ima specifičen značaj (Chargaffovo pravilo).
Linearna veriga DNK, katere dolžina je odvisna od števila nukleotidov, vključenih v verigo, ima dva konca: enega imenujemo 3" konec in vsebuje prosti hidroksil, drugega pa imenujemo 5" konec in vsebuje fosforno kislinski ostanek. Vezje je polarno in ima lahko smer 5"->3" in 3"->5". Izjema je krožna DNK.
Genetsko »besedilo« DNK je sestavljeno iz kodnih »besed« – trojčkov nukleotidov, imenovanih kodoni. Odseki DNK, ki vsebujejo informacije o primarni strukturi vseh vrst RNK, se imenujejo strukturni geni.
Polinukleotidne verige DNA dosegajo velikanske velikosti, zato so v celici zapakirane na določen način.
Chargaff (1949) je med preučevanjem sestave DNK ugotovil pomembne vzorce glede vsebine posameznih baz DNK. Pomagali so razkriti sekundarno strukturo DNK. Ti vzorci se imenujejo Chargaffova pravila. Chargaffova pravila
Ta pravila kažejo, da je treba pri konstruiranju DNK upoštevati dokaj strogo ujemanje (združevanje) ne purinskih in pirimidinskih baz na splošno, ampak posebej timina z adeninom in citozina z gvaninom. Na podlagi teh pravil sta leta 1953 Watson in Crick predlagala model sekundarne strukture DNK, imenovan dvojna vijačnica (sl.). |

Sekundarna struktura DNA
Sekundarna struktura DNK je dvojna vijačnica, katere model sta leta 1953 predlagala D. Watson in F. Crick.
Predpogoji za izdelavo modela DNK
Kot rezultat začetnih analiz je veljalo, da DNK katerega koli izvora vsebuje vse štiri nukleotide v enakih molskih količinah. Vendar pa so v štiridesetih letih 20. stoletja E. Chargaff in njegovi sodelavci z analizo DNK, izolirane iz različnih organizmov, jasno pokazali, da vsebujejo dušikove baze v različnih količinskih razmerjih. Chargaff je ugotovil, da čeprav so ta razmerja enaka za DNK iz vseh celic iste vrste organizmov, se lahko DNK različnih vrst močno razlikuje v vsebnosti določenih nukleotidov. To je nakazovalo, da so lahko razlike v razmerju dušikovih baz povezane z nekakšno biološko kodo. Čeprav se je izkazalo, da je razmerje posameznih purinskih in pirimidinskih baz v različnih vzorcih DNK različno, se je pri primerjavi rezultatov testa pokazal določen vzorec: v vseh vzorcih je bilo skupno število purinov enako skupnemu številu pirimidinov (A + G = T + C), je bila količina adenina enaka količini timina (A = T), količina gvanina pa je količina citozina (G = C). DNK, izolirana iz celic sesalcev, je bila na splošno bogatejša z adeninom in timinom ter relativno revnejša z gvaninom in citozinom, medtem ko je bila DNK iz bakterij bogatejša z gvaninom in citozinom ter relativno revnejša z adeninom in timinom. Ti podatki so bili pomemben del dejanskega gradiva, na podlagi katerega je bil kasneje zgrajen Watson-Crickov model strukture DNK.
Drug pomemben posreden pokazatelj možne strukture DNK so bili podatki L. Paulinga o zgradbi beljakovinskih molekul. Pauling je pokazal, da je možnih več različnih stabilnih konfiguracij aminokislinske verige v proteinski molekuli. Ena pogosta konfiguracija peptidne verige, α-vijačnica, je pravilna spiralna struktura. S to strukturo je možna tvorba vodikovih vezi med aminokislinami, ki se nahajajo na sosednjih obratih verige. Pauling je leta 1950 opisal α-vijačno konfiguracijo polipeptidne verige in predlagal, da imajo molekule DNA verjetno spiralno strukturo, ki jo držijo vodikove vezi.
Najdragocenejše podatke o strukturi molekule DNA pa so dali rezultati rentgenske difrakcijske analize. Rentgenski žarki, ki prehajajo skozi kristal DNA, so podvrženi difrakciji, to pomeni, da se odklonijo v določenih smereh. Stopnja in narava odklona žarkov sta odvisna od strukture samih molekul. Rentgenski difrakcijski vzorec (slika 3) daje izkušenemu očesu številne posredne indikacije glede strukture molekul preučevane snovi. Analiza rentgenskih difrakcijskih vzorcev DNK je privedla do zaključka, da so dušikove baze (ki imajo ravno obliko) razporejene kot kup plošč. Rentgenski difrakcijski vzorci so razkrili tri glavne periode v strukturi kristalne DNK: 0,34, 2 in 3,4 nm.

Watson-Crickov model DNK
Watson in Crick je začel graditi fizične modele posameznih komponent molekule DNK v določenem merilu in jih »prilagajati« drug drugemu tako, da nastali sistem ustreza različnim eksperimentalnim podatkom. [pokaži] .
Že prej je bilo znano, da so sosednji nukleotidi v verigi DNA povezani s fosfodiestrskimi mostovi, ki povezujejo 5"-ogljikov atom deoksiriboze enega nukleotida s 3"-ogljikovim atomom deoksiriboze naslednjega nukleotida. Watson in Crick nista dvomila, da obdobje 0,34 nm ustreza razdalji med zaporednimi nukleotidi v verigi DNA. Nadalje bi lahko domnevali, da obdobje 2 nm ustreza debelini verige. In da bi pojasnili, kateri dejanski strukturi ustreza obdobje 3,4 nm, sta Watson in Crick, pa tudi Pauling prej, predlagala, da je veriga zavita v obliki spirale (ali, natančneje, tvori vijačno črto, saj spiralo v strogem pomenu te besede dobimo, ko tuljave v prostoru tvorijo stožčasto in ne valjasto površino). Potem bo obdobje 3,4 nm ustrezalo razdalji med zaporednimi obrati te vijačnice. Takšna spirala je lahko zelo gosta ali nekoliko raztegnjena, to pomeni, da so njeni zavoji ravni ali strmi. Ker je perioda 3,4 nm natanko 10-kratna razdalja med zaporednimi nukleotidi (0,34 nm), je jasno, da vsak popoln zavoj vijačnice vsebuje 10 nukleotidov. Iz teh podatkov sta Watson in Crick lahko izračunala gostoto polinukleotidne verige, zvite v vijačnico s premerom 2 nm, z razdaljo med obrati 3,4 nm. Izkazalo se je, da bi imela takšna veriga gostoto, ki bi bila za polovico manjša od dejanske gostote DNK, ki je bila že znana. Moral sem domnevati, da je molekula DNK sestavljena iz dveh verig – da gre za dvojno vijačnico nukleotidov.
Naslednja naloga je bila seveda razjasniti prostorska razmerja med obema verigama, ki tvorita dvojno vijačnico. Po preizkusu številnih možnosti za razporeditev verig na svojem fizičnem modelu sta Watson in Crick ugotovila, da se vsi razpoložljivi podatki najbolje ujemajo z možnostjo, pri kateri gresta dve polinukleotidni vijačnici v nasprotnih smereh; v tem primeru verige, sestavljene iz ostankov sladkorja in fosfata, tvorijo površino dvojne vijačnice, znotraj pa se nahajajo purini in pirimidini. Baze, ki se nahajajo nasproti druge, pripadajo dvema verigama, so v parih povezane z vodikovimi vezmi; Prav te vodikove vezi držijo verige skupaj in tako fiksirajo celotno konfiguracijo molekule.
Dvojno vijačnico DNK si lahko predstavljamo kot vrvno lestev, ki je spiralno zavita, tako da njene prečke ostanejo vodoravne. Nato bosta dve vzdolžni vrvi ustrezali verigam sladkornih in fosfatnih ostankov, prečke pa bodo ustrezale parom dušikovih baz, povezanih z vodikovimi vezmi.
Kot rezultat nadaljnjega preučevanja možnih modelov sta Watson in Crick ugotovila, da mora biti vsaka "prečka" sestavljena iz enega purina in enega pirimidina; pri periodi 2 nm (kar ustreza premeru dvojne vijačnice) ne bi bilo dovolj prostora za dva purina in oba pirimidina ne bi mogla biti dovolj blizu drug drugemu, da bi tvorila ustrezne vodikove vezi. Poglobljena študija podrobnega modela je pokazala, da adenina in citozina, čeprav tvorita kombinacijo primerne velikosti, še vedno ni mogoče postaviti tako, da bi se med njima oblikovale vodikove vezi. Podobna poročila so prisilila v izključitev kombinacije gvanin - timin, medtem ko sta se kombinaciji adenin - timin in gvanin - citozin izkazali za povsem sprejemljivi. Narava vodikovih vezi je taka, da adenin tvori par s timinom, gvanin pa s citozinom. Ta zamisel o specifičnem združevanju baz je omogočila razlago "Chargaffovega pravila", po katerem je v kateri koli molekuli DNA količina adenina vedno enaka vsebnosti timina, količina gvanina pa je vedno enaka količini citozina. Med adeninom in timinom se tvorita dve vodikovi vezi, med gvaninom in citozinom pa tri. Zaradi te specifičnosti tvorba vodikovih vezi proti vsakemu adeninu v eni verigi povzroči nastanek timina na drugi; na enak način je lahko le citozin nasproti vsakemu gvaninu. Tako so verige med seboj komplementarne, to pomeni, da zaporedje nukleotidov v eni verigi enolično določa njihovo zaporedje v drugi. Dve verigi potekata v nasprotnih smereh in njuni končni fosfatni skupini sta na nasprotnih koncih dvojne vijačnice.
Kot rezultat svojih raziskav sta leta 1953 Watson in Crick predlagala model strukture molekule DNA (slika 3), ki ostaja pomemben še danes. Po modelu je molekula DNA sestavljena iz dveh komplementarnih polinukleotidnih verig. Vsaka veriga DNK je polinukleotid, sestavljen iz več deset tisoč nukleotidov. V njem sosednji nukleotidi tvorijo pravilno pentozo-fosfatno ogrodje zaradi povezave ostanka fosforne kisline in močne deoksiriboze. kovalentna vez. Dušikove baze ene polinukleotidne verige so razporejene v strogo določenem vrstnem redu nasproti dušikovih baz druge. Menjava dušikovih baz v polinukleotidni verigi je nepravilna.
Razporeditev dušikovih baz v verigi DNA je komplementarna (iz grškega "komplementa" - dodajanje), tj. Timin (T) je vedno proti adeninu (A), le citozin (C) pa proti gvaninu (G). To je razloženo z dejstvom, da A in T ter G in C strogo ustrezata drug drugemu, tj. dopolnjujeta drug drugega. Ta korespondenca je dana kemijska struktura baze, ki omogočajo tvorbo vodikovih vezi v paru purin in pirimidin. Med A in T sta dve povezavi, med G in C pa tri. Te vezi zagotavljajo delno stabilizacijo molekule DNA v prostoru. Stabilnost dvojne vijačnice je premosorazmerna s številom vezi G≡C, ki so bolj stabilne v primerjavi z vezmi A=T.
Znano zaporedje razporeditve nukleotidov v eni verigi DNK omogoča, da po principu komplementarnosti vzpostavimo nukleotide druge verige.
Poleg tega je bilo ugotovljeno, da dušikove baze z aromatično strukturo v vodna raztopina se nahajajo drug nad drugim in tvorijo kup kovancev. Ta postopek oblikovanja nizov organske molekule imenovano zlaganje. Polinukleotidne verige molekule DNK obravnavanega modela Watson-Crick imajo podobno fizikalno-kemijsko stanje, njihove dušikove baze so razporejene v obliki kupa kovancev, med ravninami katerih nastanejo van der Waalsove interakcije (interakcije zlaganja).
Vodikove vezi med komplementarnimi bazami (vodoravno) in interakcija zlaganja med ravninami baz v polinukleotidni verigi zaradi van der Waalsovih sil (navpično) zagotavlja molekuli DNA dodatno stabilizacijo v prostoru.
Sladkorfosfatna hrbtenica obeh verig je obrnjena navzven, baze pa navznoter, druga proti drugi. Smer verig v DNK je antiparalelna (ena od njih ima smer 5"->3", druga - 3"->5", tj. 3" konec ene verige se nahaja nasproti 5" konca verige. drugega.). Verige tvorijo desnosučne spirale s skupno osjo. En zavoj vijačnice je 10 nukleotidov, velikost zavoja je 3,4 nm, višina vsakega nukleotida je 0,34 nm, premer vijačnice je 2,0 nm. Kot posledica rotacije ene verige okoli druge nastaneta glavni utor (premera približno 20 Å) in manjši utor (premera približno 12 Å) dvojne vijačnice DNA. To obliko Watson-Crickove dvojne vijačnice so kasneje poimenovali B-oblika. V celicah DNK običajno obstaja v obliki B, ki je najbolj stabilna.
Funkcije DNK
Predlagani model je marsikaj pojasnil biološke lastnosti deoksiribonukleinska kislina, vključno s shranjevanjem genetskih informacij in raznolikosti genov, ki jo zagotavljajo najrazličnejše zaporedne kombinacije 4 nukleotidov in dejstvo obstoja genetske kode, sposobnost samoreprodukcije in prenosa genetskih informacij, ki jih zagotavlja replikacija proces in implementacija genetske informacije v obliki proteinov, kot tudi vseh drugih spojin, ki nastanejo s pomočjo encimskih proteinov.
Osnovne funkcije DNK.
- DNK je nosilec genetske informacije, ki je zagotovljena z dejstvom obstoja genetske kode.
- Razmnoževanje in prenos genetskih informacij med generacijami celic in organizmov. To funkcionalnost zagotavlja proces replikacije.
- Implementacija genetskih informacij v obliki beljakovin, pa tudi vseh drugih spojin, ki nastanejo s pomočjo encimskih beljakovin. To funkcijo zagotavljata procesa transkripcije in prevajanja.

Oblike organizacije dvoverižne DNK
DNK lahko tvori več vrst dvojnih vijačnic (slika 4). Trenutno je znanih že šest oblik (od A do E in Z-forma).
Strukturne oblike DNK so, kot je ugotovila Rosalind Franklin, odvisne od nasičenosti molekule nukleinske kisline z vodo. V študijah DNA vlaken z rentgensko difrakcijsko analizo je bilo dokazano, da je rentgenski vzorec močno odvisen od relativne vlažnosti, pri kateri stopnji vodne nasičenosti tega vlakna poteka poskus. Če je bilo vlakno dovolj nasičeno z vodo, je bila pridobljena ena radiografija. Po sušenju se je pojavil popolnoma drugačen rentgenski vzorec, zelo drugačen od rentgenskega vzorca vlaken z visoko vlažnostjo.
Molekulo DNK z visoko vlažnostjo imenujemo B-oblika. V fizioloških pogojih (nizka koncentracija soli, visoka stopnja hidracije) prevladuje strukturni tip DNK je B-oblika (osnovna oblika dvoverižne DNK – Watson-Crickov model). Korak vijačnice takšne molekule je 3,4 nm. Na zavoj je 10 komplementarnih parov v obliki zvitih nizov "kovancev" - dušikovih baz. Skupine držijo skupaj vodikove vezi med dvema nasprotujočima si "kovancema" skladov, "ovita" pa sta dva trakova fosfodiestrskega ogrodja, zvita v desnosučno vijačnico. Ravnine dušikovih baz so pravokotne na os vijačnice. Sosednji komplementarni pari so zasukani drug glede na drugega za 36°. Premer vijačnice je 20 Å, pri čemer purinski nukleotid zavzema 12 Å, pirimidinski nukleotid pa 8 Å.
Molekula DNK z nižjo vlažnostjo se imenuje A-oblika. A-oblika nastane v pogojih manj visoke hidracije in pri višji vsebnosti Na + ali K + ionov. Ta širša desnosučna konformacija ima 11 parov dušikovih baz na zavoj. Ravnine dušikovih baz so bolj nagnjene proti osi vijačnice, od normale na os vijačnice so odklonjene za 20°. To pomeni prisotnost notranje praznine s premerom 5Å. Razdalja med sosednjimi nukleotidi je 0,23 nm, dolžina zavoja je 2,5 nm, premer vijačnice pa 2,3 nm.
Sprva je veljalo, da je oblika DNK A manj pomembna. Vendar je pozneje postalo jasno, da ima A-oblika DNK, tako kot B-oblika, ogromen biološki pomen. Vijačnica RNA-DNA v kompleksu predloga-primer ima A-obliko, prav tako vijačnica RNA-RNA in lasne strukture RNA (2'-hidroksilna skupina riboze preprečuje molekulam RNA, da tvorijo B-obliko). A-oblika DNK se nahaja v sporah. Ugotovljeno je bilo, da je A-oblika DNK 10-krat bolj odporna na UV žarke kot B-oblika.
A-oblika in B-oblika se imenujeta kanonični obliki DNK.
Obrazci C-E prav tako desnosučne, je njihov nastanek mogoče opazovati le v posebnih poskusih in očitno ne obstajajo in vivo. C oblika DNA ima podobno strukturo kot B DNA. Število baznih parov na zavoj je 9,33, dolžina zavoja vijačnice je 3,1 nm. Osnovni pari so nagnjeni pod kotom 8 stopinj glede na pravokotni položaj na os. Žlebovi so po velikosti podobni utorom B-DNA. V tem primeru je glavni utor nekoliko plitvejši, pomožni utor pa globlji. Naravni in sintetični polinukleotidi DNA se lahko spremenijo v C-obliko.
| Tabela 1. Značilnosti nekaterih vrst struktur DNK | |||
| Spiralni tip | A | B | Z |
| Spiralni korak | 0,32 nm | 3,38 nm | 4,46 nm |
| Spiralni zasuk | Prav | Prav | levo |
| Število baznih parov na zavoj | 11 | 10 | 12 |
| Razdalja med osnovnimi ravninami | 0,256 nm | 0,338 nm | 0,371 nm |
| Konformacija glikozidne vezi | proti | proti | anti-C greh-G |
| Konformacija furanoznega obroča | C3"-endo | C2"-endo | C3"-endo-G C2"-endo-C |
| Širina utora, majhna/velika | 1,11/0,22 nm | 0,57/1,17 nm | 0,2/0,88 nm |
| Globina utora, majhna/velika | 0,26/1,30 nm | 0,82/0,85 nm | 1,38/0,37 nm |
| Premer spirale | 2,3 nm | 2,0 nm | 1,8 nm |
Strukturni elementi DNK
(nekanonične strukture DNK)
Strukturni elementi DNK vključujejo nenavadne strukture, omejene z nekaterimi posebnimi zaporedji:
|

DNK v obliki črke Z je bil odkrit leta 1979 med preučevanjem heksanukleotida d(CG)3 -. Odkrili so ga profesor MIT Alexander Rich in njegovi kolegi. Z-oblika je postala eden najpomembnejših strukturnih elementov DNA zaradi dejstva, da so njeno tvorbo opazili v regijah DNA, kjer se purini izmenjujejo s pirimidini (na primer 5'-GCGCGC-3'), ali v ponovitvah 5' -CGCGCG-3', ki vsebuje metiliran citozin. Bistven pogoj za nastanek in stabilizacijo Z-DNA je bila prisotnost purinskih nukleotidov v konformaciji sin, ki se izmenjujejo s pirimidinskimi bazami v antikonformaciji.
Naravne molekule DNK večinoma obstajajo v desnosučni B-obliki, razen če vsebujejo zaporedja, kot je (CG)n. Če pa so takšna zaporedja del DNK, potem se lahko ti odseki, ko se spremeni ionska moč raztopine ali kationov, ki nevtralizirajo negativni naboj na fosfodiestrskem ogrodju, spremenijo v Z-obliko, medtem ko se drugi odseki DNK v veriga ostane v klasični B-obliki. Možnost takšnega prehoda kaže, da sta obe verigi v dvojni vijačnici DNK v dinamičnem stanju in se lahko odvijata relativno druga proti drugi, premikajoč se iz desne oblike v levo in obratno. Biološke posledice takšne labilnosti, ki omogoča konformacijske preobrazbe strukture DNA, še niso povsem razumljene. Menijo, da deli Z-DNA igrajo določeno vlogo pri uravnavanju izražanja določenih genov in sodelujejo pri genetski rekombinaciji.
Z-oblika DNK je levosučna dvojna vijačnica, v kateri se fosfodiestersko ogrodje nahaja v cikcakastem vzorcu vzdolž osi molekule. Od tod tudi ime molekule (cikcak)-DNK. Z-DNK je najmanj zavita (12 baznih parov na zavoj) in najtanjša DNK, kar jih poznamo v naravi. Razdalja med sosednjima nukleotidoma je 0,38 nm, dolžina zavoja 4,56 nm, premer Z-DNA pa 1,8 nm. Poleg tega se videz te molekule DNK odlikuje po prisotnosti enega utora.
Z obliko DNK so našli v prokariontskih in evkariontskih celicah. Zdaj so bila pridobljena protitelesa, ki lahko ločijo Z-obliko od B-oblike DNA. Ta protitelesa se vežejo na specifične predele velikanskih kromosomov celic žleze slinavke Drosophila (Dr. melanogaster). Reakcijo vezave je enostavno spremljati zaradi nenavadne strukture teh kromosomov, v katerih so gostejše regije (diski) v nasprotju z manj gostimi regijami (interdiski). Regije Z-DNA se nahajajo v interdiskih. Iz tega sledi, da Z-oblika dejansko obstaja v naravnih razmerah, čeprav velikosti posameznih odsekov Z-oblike še niso znane.

(inverterji) so najbolj znana in pogosto pojavljajoča se bazna zaporedja v DNK. Palindrom je beseda ali fraza, ki se bere enako od leve proti desni in obratno. Primeri takšnih besed ali besednih zvez so: KOČA, KOZAČ, POPLAVA IN VRTNICA JE PADALA NA AZORJEVO ŠAPO. Ko se nanaša na odseke DNK, ta izraz (palindrom) pomeni enako menjavo nukleotidov vzdolž verige od desne proti levi in od leve proti desni (kot črke v besedi "koča" itd.).
Za palindrom je značilna prisotnost obrnjenih ponovitev baznih zaporedij, ki imajo simetrijo drugega reda glede na dve verigi DNA. Takšna zaporedja so iz očitnih razlogov samokomplementarna in težijo k oblikovanju lasnih ali križnih struktur (sl.). Lasnice pomagajo regulatornim beljakovinam prepoznati, kje je kopirano genetsko besedilo kromosomske DNK.
Če je na isti verigi DNK prisotna obrnjena ponovitev, se zaporedje imenuje zrcalna ponovitev. Zrcalne ponovitve nimajo lastnosti samokomplementarnosti in zato niso sposobne oblikovati lasnih ali križnih struktur. Zaporedja te vrste najdemo v skoraj vseh velikih molekulah DNK in lahko segajo od le nekaj baznih parov do nekaj tisoč baznih parov.
Prisotnost palindromov v obliki križnih struktur v evkariontskih celicah ni bila dokazana, čeprav je bilo določeno število križnih struktur odkritih in vivo v celicah E. coli. Prisotnost samokomplementarnih zaporedij v RNA ali enoverižni DNA je glavni razlog za zvijanje verige nukleinske kisline v raztopinah v določeno prostorsko strukturo, za katero je značilno nastajanje številnih "lasnic".

H-oblika DNK je vijačnica, ki jo tvorijo tri verige DNK – trojna vijačnica DNK. Je kompleks Watson-Crickove dvojne vijačnice s tretjo enoverižno verigo DNA, ki se prilega njenemu glavnemu utoru in tvori tako imenovani Hoogsteenov par.
Tvorba takega tripleksa se pojavi kot posledica prepogibanja dvojne vijačnice DNA tako, da polovica njenega dela ostane v obliki dvojne vijačnice, druga polovica pa se loči. V tem primeru ena od nepovezanih vijačnic tvori novo strukturo s prvo polovico dvojne vijačnice - trojno vijačnico, druga pa se izkaže za nestrukturirano, v obliki enoverižnega odseka. Značilnost tega strukturnega prehoda je njegova močna odvisnost od pH medija, katerega protoni stabilizirajo novo strukturo. Zaradi te lastnosti novo strukturo se je imenovala H-oblika DNA, katere nastanek so odkrili v superzvitih plazmidih, ki vsebujejo homopurin-homopirimidinske regije, ki so zrcalna ponovitev.
V nadaljnjih študijah je bilo ugotovljeno, da je možno izvesti strukturni prehod nekaterih homopurin-homopirimidinskih dvoverižnih polinukleotidov s tvorbo triverižne strukture, ki vsebuje:
- ena homopurinska in dve homopirimidinska veriga ( Py-Pu-Py tripleks) [Hoogsteenova interakcija].
Sestavni bloki tripleksa Py-Pu-Py so kanonične izomorfne triade CGC+ in TAT. Stabilizacija tripleksa zahteva protonacijo triade CGC+, zato so ti tripleksi odvisni od pH raztopine.
- ena homopirimidinska in dve homopurinska veriga ( Py-Pu-Pu tripleks) [inverzna Hoogsteenova interakcija].
Sestavni bloki tripleksa Py-Pu-Pu so kanonične izomorfne triade CGG in TAA. Bistvena lastnost tripleksov Py-Pu-Pu je odvisnost njihove stabilnosti od prisotnosti dvojno nabitih ionov, za stabilizacijo tripleksov različnih zaporedij pa so potrebni različni ioni. Ker tvorba tripleksov Py-Pu-Pu ne zahteva protonacije njihovih sestavnih nukleotidov, lahko takšni tripleksi obstajajo pri nevtralnem pH.
Opomba: direktne in reverzne Hoogsteenove interakcije so razložene s simetrijo 1-metiltimina: rotacija za 180° povzroči, da atom O2 prevzame mesto atoma O4, medtem ko se sistem vodikovih vezi ohrani.
Znani sta dve vrsti trojnih vijačnic:
- vzporedne trojne vijačnice, v katerih polarnost tretje verige sovpada s polarnostjo homopurinske verige Watson-Crickovega dupleksa
- antiparalelne trojne vijačnice, v katerih sta polarnosti tretje in homopurinske verige nasprotni.

G-kvadrupleks- 4-verižna DNA. Ta struktura nastane, če obstajajo štirje gvanini, ki tvorijo tako imenovani G-kvadrupleks - krožni ples štirih gvaninov.
Prvi namigi o možnosti oblikovanja takšnih struktur so bili prejeti že dolgo pred prelomnim delom Watsona in Cricka - leta 1910. Nato je nemški kemik Ivar Bang odkril, da ena od komponent DNK – gvanozinska kislina – pri visokih koncentracijah tvori gele, medtem ko druge komponente DNK te lastnosti nimajo.
Leta 1962 je bilo z metodo rentgenske difrakcije mogoče ugotoviti celično strukturo tega gela. Izkazalo se je, da je sestavljen iz štirih ostankov gvanina, ki se med seboj povezujejo v krog in tvorijo značilen kvadrat. V središču je vez podprta s kovinskim ionom (Na, K, Mg). Enake strukture se lahko tvorijo v DNK, če vsebuje veliko gvanina. Ti ploski kvadrati (G-kvarteti) so zloženi tako, da tvorijo precej stabilne, goste strukture (G-kvadrupleksi).
Štiri ločene verige DNK so lahko vtkane v štiriverižne komplekse, vendar je to precej izjema. Pogosteje je posamezna veriga nukleinske kisline preprosto povezana v vozel, ki tvori značilne zadebelitve (na primer na koncih kromosomov), ali pa dvoverižna DNK na nekem z gvaninom bogatem območju tvori lokalni kvadrupleks.
Najbolj je bil raziskan obstoj kvadrupleksov na koncih kromosomov – na telomerah in v tumorskih promotorjih.
Vendar pa popolna slika o lokalizaciji takšne DNK v človeških kromosomih še vedno ni znana.
- Vse te nenavadne strukture DNK v linearni obliki so nestabilne v primerjavi z B-obliko DNK. Vendar DNK pogosto obstaja v krožni obliki topološke napetosti, ko ima tako imenovano superzvijanje. V teh pogojih se zlahka oblikujejo nekanonične strukture DNK: Z-oblike, "križi" in "lasnice", H-oblike, gvaninski kvadrupleksi in i-motiv.
- Superzvita oblika – opažena, ko se sprosti iz celičnega jedra, ne da bi poškodovala ogrodje pentozofosfata. Ima obliko super zvitih zaprtih obročev. V superzvitem stanju je dvojna vijačnica DNA vsaj enkrat "zasukana vase", to pomeni, da vsebuje vsaj en superturn (ima obliko osmice).
- Sproščeno stanje DNK – opazno z enim samim prelomom (prelom ene verige). V tem primeru superzvitki izginejo in DNK prevzame obliko sklenjenega obroča.
Vse te tri oblike DNK zlahka ločimo z gelsko elektroforezo.
Terciarna struktura DNK Terciarna struktura DNK
Skoraj vsa DNK evkariontov se nahaja v kromosomih jeder; le majhna količina je v mitohondrijih, v rastlinah pa v plastidih. Glavna snov kromosomov evkariontskih celic (vključno s človeškimi kromosomi) je kromatin, sestavljen iz dvoverižne DNA, histonskih in nehistonskih proteinov.
Proteini histonskega kromatina
Histoni so enostavni proteini, ki tvorijo do 50 % kromatina. V vseh proučevanih živalskih in rastlinskih celicah je bilo najdenih pet glavnih razredov histonov: H1, H2A, H2B, H3, H4, ki se razlikujejo po velikosti, aminokislinska sestava in količino naboja (vedno pozitivno).
Histon H1 pri sesalcih je sestavljen iz ene same polipeptidne verige, ki vsebuje približno 215 aminokislin; velikosti drugih histonov se razlikujejo od 100 do 135 aminokislin. Vsi so spiralizirani in zviti v globulo s premerom približno 2,5 nm ter vsebujejo nenavadno veliko količino pozitivno nabitih aminokislin lizina in arginina. Histoni so lahko acetilirani, metilirani, fosforilirani, poli(ADP)-ribozilirani, histona H2A in H2B pa sta kovalentno povezana z ubikvitinom. Vloga takšnih modifikacij pri oblikovanju strukture in opravljanju funkcij histonov še ni povsem pojasnjena. Predpostavlja se, da je to njihova sposobnost interakcije z DNK in zagotavljanja enega od mehanizmov za uravnavanje delovanja genov.
Histoni medsebojno delujejo z DNK predvsem preko ionske vezi(solni mostovi), ki nastanejo med negativno nabitimi fosfatnimi skupinami DNK in pozitivno nabitimi lizinskimi in argininskimi ostanki histonov.
Nehistonski kromatinski proteini
Nehistonski proteini so za razliko od histonov zelo raznoliki. Izoliranih je bilo do 590 različnih frakcij nehistonskih proteinov, ki vežejo DNA. Imenujemo jih tudi kisli proteini, saj v njihovi strukturi prevladujejo kisle aminokisline(so polianioni). Raznolikost nehistonskih proteinov je povezana s specifično regulacijo aktivnosti kromatina. Na primer, encimi, potrebni za replikacijo in izražanje DNA, se lahko začasno vežejo na kromatin. Drugi proteini, recimo tisti, ki sodelujejo v različnih regulacijskih procesih, se na DNK vežejo le v specifičnih tkivih ali na določenih stopnjah diferenciacije. Vsak protein je komplementaren določenemu zaporedju nukleotidov DNA (mestu DNA). Ta skupina vključuje:
- družina proteinov cinkovega prsta, specifičnih za mesto. Vsak »cinkov prst« prepozna določeno mesto, sestavljeno iz 5 nukleotidnih parov.
- družina za mesto specifičnih proteinov – homodimerov. Fragment takega proteina v stiku z DNA ima strukturo vijačnica-obrat-vijačnica.
- gelski proteini z visoko mobilnostjo (HMG proteini) so skupina strukturnih in regulatornih proteinov, ki so stalno povezani s kromatinom. Imajo molekulsko maso manjšo od 30 kDa in zanje je značilna visoka vsebnost nabitih aminokislin. Zaradi nizke molekulske mase imajo HMG proteini visoko mobilnost med elektroforezo v poliakrilamidnem gelu.
- replikacijski, transkripcijski in popravljalni encimi.
S sodelovanjem strukturnih, regulatornih proteinov in encimov, ki sodelujejo pri sintezi DNK in RNK, se nit nukleosoma pretvori v visoko kondenziran kompleks proteinov in DNK. Nastala struktura je 10.000-krat krajša od prvotne molekule DNK.

Kromatin
Kromatin je kompleks beljakovin z jedrno DNA in anorganske snovi. Glavnina kromatina je neaktivna. Vsebuje tesno zapakirano, zgoščeno DNK. To je heterokromatin. Obstaja konstitutivni, genetsko neaktiven kromatin (satelitska DNK), sestavljen iz neizraženih regij, in fakultativni - neaktiven v številnih generacijah, vendar pod določenimi pogoji sposoben izražanja.
Aktivni kromatin (evkromatin) je nekondenziran, tj. zapakirano manj tesno. V različnih celicah se njegova vsebnost giblje od 2 do 11%. V možganskih celicah ga je največ - 10-11%, v jetrnih celicah - 3-4 in ledvičnih celicah - 2-3%. Opažena je aktivna transkripcija evhromatina. Poleg tega njegova strukturna organizacija omogoča uporabo iste genetske informacije DNK, ki je inherentna te vrste telesu, drugače v specializiranih celicah.
Pod elektronskim mikroskopom je slika kromatina podobna kroglicam: sferične odebelitve velikosti približno 10 nm, ločene z nitastimi mostički. Te sferične odebelitve imenujemo nukleosomi. Nukleosom je strukturna enota kromatina. Vsak nukleosom vsebuje 146-bp superzvit segment DNA, navit v 1,75 levih zavojev na nukleosomsko jedro. Nukleosomsko jedro je histonski oktamer, sestavljen iz histonov H2A, H2B, H3 in H4, dveh molekul vsake vrste (slika 9), ki izgleda kot disk s premerom 11 nm in debelino 5,7 nm. Peti histon, H1, ni del nukleosomskega jedra in ni vključen v proces navijanja DNA na histonski oktamer. V stiku je z DNK na mestih, kjer dvojna vijačnica vstopa in izstopa iz nukleosomskega jedra. To so medjedrni (povezovalni) odseki DNA, katerih dolžina se glede na tip celice spreminja od 40 do 50 nukleotidnih parov. Posledično se spreminja tudi dolžina fragmenta DNA, ki je vključen v nukleosome (od 186 do 196 nukleotidnih parov).
Nukleosomi vsebujejo približno 90 % DNK, ostalo so povezovalci. Menijo, da so nukleozomi fragmenti "tihega" kromatina in da je povezovalec aktiven. Vendar pa se lahko nukleosomi razvijejo in postanejo linearni. Razviti nukleosomi so že aktivni kromatin. To jasno dokazuje odvisnost funkcije od strukture. Lahko domnevamo, da več kromatina vsebuje globularni nukleosom, manj je aktiven. Očitno je v različnih celicah neenakomeren delež kromatina v mirovanju povezan s številom takšnih nukleosomov.
Na elektronskih mikroskopskih fotografijah lahko kromatin, odvisno od pogojev izolacije in stopnje raztezanja, izgleda ne le kot dolga nit z zadebelitvami - "kroglicami" nukleosomov, temveč tudi kot krajša in gostejša fibrila (vlakno) s premerom 30 nm, katerega tvorbo opazimo med interakcijo histona H1, vezanega na povezovalno regijo DNA in histona H3, kar vodi do dodatnega zvijanja vijačnice šestih nukleosomov na obrat, da se tvori solenoid s premerom 30 nm. V tem primeru lahko protein histon moti prepisovanje številnih genov in tako uravnava njihovo aktivnost.
Kot rezultat zgoraj opisanih interakcij DNA s histoni se segment dvojne vijačnice DNA s 186 baznimi pari s povprečnim premerom 2 nm in dolžino 57 nm pretvori v vijačnico s premerom 10 nm in dolžine 5 nm. Ko se ta vijačnica naknadno stisne v vlakno s premerom 30 nm, se stopnja kondenzacije poveča še šestkrat.
Navsezadnje pakiranje dupleksa DNK s petimi histoni povzroči 50-kratno kondenzacijo DNK. Vendar pa še tako visoka stopnja kondenzacije ne more razložiti skoraj 50.000 - 100.000-kratne kompaktnosti DNK v metafaznem kromosomu. Na žalost podrobnosti o nadaljnjem pakiranju kromatina do metafaznega kromosoma še niso znane, zato lahko razmišljamo le o splošne značilnosti ta proces.
Stopnje zbijanja DNK v kromosomih
Vsaka molekula DNK je zapakirana v ločen kromosom. Človeške diploidne celice vsebujejo 46 kromosomov, ki se nahajajo v celičnem jedru. Skupna dolžina DNK vseh kromosomov v celici je 1,74 m, vendar je premer jedra, v katerem so zapakirani kromosomi, milijonkrat manjši. Tako kompaktno pakiranje DNK v kromosomih in kromosomov v celičnem jedru zagotavljajo različni histonski in nehistonski proteini, ki v določenem zaporedju interagirajo z DNK (glej zgoraj). Zgoščevanje DNK v kromosomih omogoča zmanjšanje njenih linearnih dimenzij za približno 10.000-krat - približno od 5 cm do 5 mikronov. Obstaja več stopenj zbijanja (slika 10).


- Dvojna vijačnica DNA je negativno nabita molekula s premerom 2 nm in dolžino nekaj cm.
- ravni nukleosomov- kromatin izgleda v elektronskem mikroskopu kot veriga "kroglic" - nukleosomov - "na niti". Nukleosom je univerzalna strukturna enota, ki jo najdemo v evhromatinu in heterokromatinu, v interfaznem jedru in metafaznih kromosomih.
Nukleosomsko raven zbijanja zagotavljajo posebni proteini - histoni. Osem pozitivno nabitih histonskih domen tvori jedro nukleosoma, okoli katerega je ovita negativno nabita molekula DNA. To daje 7-kratno skrajšanje, premer pa se poveča z 2 na 11 nm.
- nivo solenoida
Za solenoidno raven organizacije kromosomov je značilno zvijanje nukleosomskega filamenta in tvorba debelejših fibril s premerom 20-35 nm - solenoidov ali superbidov. Korak solenoida je približno 6-10 nukleosomov na obrat. Solenoidno pakiranje velja za bolj verjetno kot superbidno pakiranje, po katerem je kromatinska fibrila s premerom 20-35 nm veriga granul ali superbidov, od katerih je vsak sestavljen iz osmih nukleosomov. Na ravni solenoida se linearna velikost DNK zmanjša za 6-10 krat, premer se poveča na 30 nm.
- raven zanke
Nivo zanke zagotavljajo proteini, ki vežejo DNK, ki niso specifični za histonsko mesto in ki prepoznajo in se vežejo na specifične sekvence DNK, pri čemer tvorijo zanke velikosti približno 30-300 kb. Zanka zagotavlja izražanje genov, tj. zanka ni samo strukturna, ampak tudi funkcionalna tvorba. Skrajšanje na tej ravni se pojavi 20-30-krat. Premer se poveča na 300 nm. V citoloških pripravkih lahko opazimo strukture v obliki zank, kot so "ščetke svetilke" v jajčnih celicah dvoživk. Zdi se, da so te zanke superzvite in predstavljajo domene DNA, ki verjetno ustrezajo enotam transkripcije in replikacije kromatina. Specifični proteini fiksirajo baze zank in po možnosti nekatere njihove notranje dele. Organizacija domene v obliki zanke spodbuja zvijanje kromatina v metafaznih kromosomih v vijačne strukture višjih redov.
- ravni domene
Domenska raven organizacije kromosomov ni bila dovolj raziskana. Na tej ravni opazimo tvorbo zančnih domen - strukture niti (fibrile) debeline 25-30 nm, ki vsebujejo 60% beljakovin, 35% DNA in 5% RNA, so praktično nevidne v vseh fazah celičnega cikla z razen mitoze in so nekoliko naključno porazdeljeni po celičnem jedru. V citoloških pripravkih lahko opazimo strukture v obliki zank, kot so "ščetke svetilke" v jajčnih celicah dvoživk.
Domene zank so na svoji osnovi pritrjene na intranuklearni proteinski matriks v tako imenovanih vgrajenih pritrdilnih mestih, ki jih pogosto imenujemo zaporedja MAR/SAR (MAR, iz angleške matrix associated region; SAR, iz angleške scaffold attachment regions) - Fragmenti DNA z več sto dolgimi baznimi pari, za katere je značilna visoka vsebnost (>65 %) A/T nukleotidnih parov. Zdi se, da ima vsaka domena en sam izvor replikacije in deluje kot avtonomna supervijačna enota. Vsaka domena zanke vsebuje veliko transkripcijskih enot, katerih delovanje je verjetno usklajeno – celotna domena je v aktivnem ali neaktivnem stanju.
Na nivoju domene se zaradi zaporednega pakiranja kromatina linearne dimenzije DNK zmanjšajo za približno 200-krat (700 nm).
- kromosomski ravni
Na kromosomski ravni pride do kondenzacije profaznega kromosoma v metafazni kromosom s stiskanjem zankalnih domen okoli aksialnega ogrodja nehistonskih proteinov. To superzvijanje spremlja fosforilacija vseh molekul H1 v celici. Posledično lahko metafazni kromosom prikažemo kot tesno zapakirane solenoidne zanke, zvite v tesno spiralo. Tipičen človeški kromosom lahko vsebuje do 2600 zank. Debelina takšne strukture doseže 1400 nm (dve kromatidi), molekula DNA pa se skrajša za 104-krat, tj. od 5 cm raztegnjene DNA do 5 µm.
Funkcije kromosomov
V interakciji z ekstrakromosomskimi mehanizmi kromosomi zagotavljajo
- shranjevanje dednih informacij
- uporabo teh informacij za ustvarjanje in vzdrževanje celične organizacije
- regulacija branja dednih informacij
- samopodvajanje genskega materiala
- prenos genskega materiala iz matične celice v hčerinske celice.
Obstajajo dokazi, da ko se aktivira regija kromatina, tj. med transkripcijo se iz njega reverzibilno odstrani najprej histon H1 in nato histonski oktet. To povzroči dekondenzacijo kromatina, sekvenčni prehod 30-nm kromatinske fibrile v 10-nm fibrilo in njegovo nadaljnje odvijanje v odseke proste DNA, t.j. izguba strukture nukleosomov.
Ekologija znanja. Znanost in odkritja: Kaj je življenje? To vprašanje je postalo gonilna sila razvoj genetike (iz grščine genetikos - »v zvezi z rojstvom, izvor«) - znanost o izvoru življenja, katere središče je že več kot 50 let molekula DNK.
Kaj je življenje? To vprašanje je postalo gonilna sila razvoja genetike (iz grščine genetikos, »v zvezi z rojstvom, izvorom«), znanosti o izvoru življenja, ki se že več kot 50 let osredotoča na molekulo DNK.
Odkritje, ki je obrnilo svet na glavo
"Pravkar smo odkrili skrivnost življenja!" - tako sta 28. februarja 1953 Francis Crick in James Watson poročala o svojem odkritju strukture DNK. Kaj je novega prinesel v znanosti o življenju? Pred tem je bilo znano, da je DNK velika molekula, v kateri so s pomočjo »štiričrkovne abecede« zapisane informacije o zgradbi in lastnostih živih bitij. Vendar je ostalo nejasno, kako se te informacije prenašajo iz generacije v generacijo in materializirajo v te iste strukture in lastnosti ter kakšna je prostorska struktura DNK.
Razkritje strukture DNK je znanstvenikom pomagalo razumeti mehanizme njenega kopiranja in materializacije. DNK je sestavljena iz dveh verig, ki sta med seboj komplementarni (komplementarni). Do kopiranja DNK pride zaradi dodajanja dodatne kopije vsaki izvirni verigi DNK, kot na matrici. Tako iz ene dvojne vijačnice DNK dobimo dve dvojni vijačnici, ki sta ji popolnoma enaki, kar je potrebno za ohranitev genetske informacije, ko se matična celica deli na dve hčerinski celici. Načelo matrike je tudi podlaga za postopno materializacijo genetske informacije: na eni od verig DNA se oblikuje komplementarna veriga druge informacijske molekule - RNA, ki služi kot matrika za sintezo beljakovin, katerih količina in kakovost določata zgradbo in lastnosti določenega organizma.
Kako pomembno je to odkritje za razumevanje skrivnosti življenja? Po eni strani poznavanje strukture DNK očitno ni dovolj za odgovor na vprašanje "kaj je življenje?" Toda po drugi strani je prav to odkritje naredilo zelo starodavno in zelo pomembno vprašanje o razmerju med potencialom in manifestom - na primeru povezave informacij o strukturah in lastnostih telesa s strukturami in lastnostmi samimi. In ne samo, da je odprlo to vprašanje, ampak tudi dalo ključ do odgovora nanj. Ta ključ je matrični princip, princip komplementarnosti.
Pot od gena do lastnosti
Kaj pomeni klasični stavek iz učbenika: »DNK je nosilec genetske informacije«? Kako so genetske informacije povezane s strukturo DNK? Kako so informacije utelešene v specifičnih lastnostih organizma? Če za izhodišče genetske informacije vzamemo strukturo DNK in nato sledimo strukturnemu modelu, potem bo pot od gena do lastnosti videti takole: vse lastnosti organizma so šifrirane v zaporedju DNK; linearna struktura določenega gena nedvoumno določa linearno strukturo njegovega ustreznega proteina, ta pa nedvoumno določa vlogo tega proteina pri nastanku določene lastnosti.
Z drugimi besedami, »DNK rodi RNK; RNA rodi beljakovine, beljakovine pa rodijo tebe in mene« (F. Crick). Če je to res, potem je za spremembo določene lastnosti (na primer za zdravljenje bolezni, ki ima genetske korenine) dovolj, da določimo ustrezno zaporedje DNK in ga popravimo.
Toda ali je res tako preprosto? Ali obstaja dovolj znanja (čeprav je nedvomno resnično in potrebno!) o strukturnih korespondencah na poti od gena do lastnosti, da bi to pot razumeli in reproducirali?
Nedavni napredek v genetiki je pokazal, da so nezadostni. Leta 2003 je Projekt človeškega genoma v celoti določil linearno strukturo človeške DNK (in mnogih drugih preprostih in kompleksnih organizmov). Kot je dejal eden od znanstvenikov: "Prebral sem vse črke, s katerimi je napisana debela knjiga, zdaj bi rad razumel besede in njihov pomen."
Izkazalo se je, da imamo ljudje v resnici približno 30.000 genov (odsekov DNK, ki kodirajo proteine), in to je le 1–3 % vse DNK! Rastlina Arabidopsis taliana in riba napihovalka imata enako število genov. Poleg tega je 99 % človeških genov enakih kot mišji geni, kar pomeni, da imamo ljudje samo 300 genov, ki jih miši nimajo. (Težko si je predstavljati, da imamo miši in tudi mi 99 % enakih lastnosti!)
Nadalje - več. Izkazalo se je, da nedvoumno razmerje med genom in beljakovino obstaja le pri bakterijah. In pri ljudeh je možno, da na podlagi enega gena nastane veliko proteinov (danes znano največje število različnih proteinov, ki jih kodira en gen, je 40.000!) in v enem proteinu nastane veliko funkcij. Izkazalo se je, da pot od potenciala do manifestacije, od genetske informacije do lastnosti nikakor ni linearna; da je vsak znak rezultat kompleksne interakcije veliko genov in njihovih beljakovinskih produktov; da sam pojem »gena« zaradi svoje dvoumnosti težko služi kot »izhodišče« na tej poti.
Interakcija struktur
Človeško telo sestavlja 10 do 14. potenca celic. Vsi imajo popolnoma enak DNK, vendar se bistveno razlikujejo po obliki, velikosti in svojih nalogah. Rešitev tega paradoksa je v selektivnem branju genetskih informacij. V vsaki celici so aktivni le tisti geni, ki so zanjo pomembni. v tem trenutku potrebno.
Selektivnost zagotavljajo posebni regulatorni geni, ki omogočajo ali prepovedujejo branje informacij iz določenega dela DNK. Delovanje gena je odvisno tudi od njegovega prostorskega okolja celično jedro. Sprememba okolja, ki jo povzroči gibanje samega gena ali katerega od njegovih sosedov, lahko spremeni njegovo aktivnost (»izklopi« ali »vklopi« gen).
Na primer, človeški genom vsebuje veliko potencialno nevarnih virusnih genov in protoonkogenov (ki lahko povzročijo rakavo degeneracijo celice). Lahko se dolgo (in vse življenje) obnašajo povsem mirno in celo delujejo v korist celice, dokler gibanje njih samih ali nekoga iz okolice ne razkrije agresivnih potencialov v teh genih. Na srečo se lahko pojavijo drugi premiki, ki bodo "upornika" pomirili ali aktivirali obrambne mehanizme.
Torej se je nosilec genetske informacije premaknil z ravni gena (določen odsek DNK) na epigenetsko (iz latinščine »zgoraj«, »nad«) raven interakcije genov med seboj in z drugimi strukturami celično jedro (99 % negenske DNK in proteinov). Recimo, da znanost razvozla mehanizem te interakcije. Bo to vodilo k odkritju skrivnosti življenja? Je življenje samo struktura? In če ne, ali naj bo iskanje odgovorov na skrivnost življenja omejeno na interakcijo struktur?
Kdo pazi na čuvaja?
Kako ena sama celica zaradi 46 delitev ne proizvede brezoblične mase 1014 celic, ampak zelo značilno telo vsakega od nas? Z zaporednim podvajanjem celice ne postanejo drugačne le same, ampak tvorijo tudi različne dele telesa pravi čas in na pravem mestu. Kaj nadzoruje organizacijo celic v času in prostoru?
Celota, ki je kvalitativno večja od enostavne vsote svojih sestavnih delov – celic. In to ni v nasprotju z dejstvom, da je organizem sestavljen iz ene celice - vprašanje je, kaj za to celico uteleša "voljo celote". Iskanje takega dejavnika urejanja je v začetku 20. stoletja privedlo do koncepta morfogenetskega polja. Njegov ustanovitelj je bil ruski znanstvenik A.G. Gurvič. Ko je Gurvich delal na teoriji polja, je bila obravnavana molekula DNK sestavni del kromosomov in mu niso pripisovali velikega pomena.
Leta 1944 je znanstvenik objavil svoje delo "Teorija biološkega polja". To leto je postalo usodno za vso genetiko in je določilo pot njenega razvoja za nekaj desetletij naprej. Pozornost znanstvenikov je bila na molekuli DNK, saj je dokazano, da igra vodilno vlogo pri prenosu dednih informacij. Leto 1953 je bilo pred vrati... Posledica tega je bila, da je znanost vso svojo pozornost usmerila v strukturo DNK, ki so jo dejansko začeli identificirati z informacijami, ki jih nosi, teorija biološkega polja pa ni bila cenjena. Toda raziskave na tem področju so se nadaljevale in vsa ta leta sta dve poti do razumevanja skrivnosti življenja potekali vzporedno ...
Prehod v novo tisočletje je spremenil razmerje moči v znanostih o življenju. Vse več znanstvenikov prihaja do zaključka, da je strukturni ključ v poznavanju živih bitij nujen, a ne zadosten; da se različni pristopi ne izključujejo, temveč tvorijo enotno pot znanstvenega raziskovanja; da sta v bistvu strukturni pristop in teorija polja komplementarna.
Spomnimo: prav domneva o komplementarnosti verig DNK je postala ključna za razvozlanje njene zgradbe, samo odkritje leta 1953 pa se je izkazalo za možno zaradi komplementarnih prizadevanj predstavnikov različnih področij znanosti – fizikov, kemikov, biologi. Morda združena znanost novega tisočletja ne bo le končno uskladila različnih znanstvenih pristopov (na primer strukturnega in terenskega), ampak se bo obrnila tudi na sadove »neznanstvene« poti razumevanja skrivnosti življenja - tisočletnega stara modrost človeštva - "neznanstvena", saj ta pot izvira iz tistih časov, ko o znanosti še ni bilo sledu.
Obrnitev k virom starodavne modrosti lahko znanosti da ključ do vrat, za katerimi se skriva skrivnost. Toda da bi se to zgodilo, tako različne načine znanje se mora nekje "presekati". Eno od teh »križišč« je lahko koncept formativnega polja (biološkega, morfogenetskega, informacijskega), ki je zraslo na podlagi moderna znanost in vračanje k pojavom, obravnavanim v starodavnih virih.
Slednji pravijo, da je človek sestavljen iz več teles ali principov, ki niso ločeni, neodvisni deli, ampak se medsebojno prepletajo in med seboj oblikujejo; da je vidno, gosto, fizično telo prevodnik, nosilec subtilnejših teles, ki se z njegovo pomočjo manifestirajo v fizični svet in komunicirati z njim; da "sestavo" - nastanek fizičnega telesa iz elementov fizične materije - nadzira najbolj "gost" teh teles, astralno telo-prototip (ravnina, matrica).
Sodobna znanost ve, da vsaka novonastala celica sodeluje pri nastajanju telesa po individualnih »navodilih« (aktivnih ali vklopljenih genih) in da imajo starši in potomci te celice lahko povsem drugačna »navodila«. Toda kaj in kako usklajuje preklapljanje posameznih "programov" za razvoj milijard celic, še ni jasno.
Teorija biološkega polja predlaga, da je koordinacija funkcija celote, ki je to isto polje, matrica ali model; da je vsaka novonastala celica s pomočjo lastnega genetskega aparata povezana v en sam »načrt« razvoja organizma, od tam prejema individualna »navodila« in jih izvaja v okviru lastnega vedenjskega programa.
Izkazalo se je, da je genetski aparat celice sestavljen iz vsaj treh funkcionalnih blokov: zaznavne "antene", "nadzorne plošče" za aktivnost genov in "izvršilnega" dela - genov, odgovornih za tvorbo specifičnih beljakovin. Ne pozabite, da geni predstavljajo le 1–2 % celotne celične DNK. V preostalih 98–99 % DNK so že našli strukture, ki pripadajo drugemu "kontrolnemu bloku". Kaj igra vlogo "antene"? Kje se zgodi "srečanje dveh svetov" - informacijskega polja in genetskih struktur, ki te informacije utelešajo v določenem fizičnem telesu?
Zakaj ne domnevamo, da ima DNK tudi vlogo antene, ki je sposobna zajemati, transformirati in prenašati signale valovne narave? To je posledica spiralne strukture »molekule življenja« (številne tehnične antene imajo obliko spirale) in njenih lastnosti, kot je sposobnost prevajanja električni tok, možnost resonančnega vzbujanja vzdolžnih vibracij pod vplivom radijskih valov, kot tudi sposobnost generiranja laserske svetlobe po predhodnem "črpanju".
Če DNK lahko dela za sprejemanje informacij, ki zagotavljajo vitalno aktivnost celic, potem ni nujno, da te informacije trajno shrani v svoji strukturi. Tako kot na primer človeški možgani za uspešno upravljanje telesnih sistemov za vzdrževanje življenja ne rabijo biti »vsebnik« uma, ampak igrajo vlogo posrednika med zavestjo in telesom: zaznavajo informacije iz ravni zavesti in jih »prevede« v jezik obvladovanja telesa.
In jasno je, zakaj če je struktura DNK (ali možganske strukture) poškodovana, fizično telo trpi. Navsezadnje vsi vedo, da če se vsaj en del televizorja pokvari, je slika na njegovem zaslonu močno popačena, in če televizorju odvzamemo anteno ali ga izklopimo iz omrežja, se na zaslonu sploh ne prikaže nič.
DNK je povezovalni člen med »modelom« fizičnega telesa in njegovim specifičnim utelešenjem. Možgani so posrednik med umom in telesom. Um povezuje življenje in obliko njegove manifestacije ter omogoča življenju, vsebovanemu v obliki, da spozna samo sebe. S pomočjo tega čudovitega orodja ima človek možnost študija svet okoli nas in v njem najti ključ do razumevanja svojega notranjega sveta. Tako se rodi enotna pot, ki vodi do spoznanja skrivnosti življenja. Kajti človek je največja izmed skrivnosti – skrivnost razmerja med zemljo in nebom. objavljeno
P.S. In ne pozabite, samo s spreminjanjem vaše porabe skupaj spreminjamo svet! © ekonet
Pridružite se nam
Po odkritju načela molekularne organizacije snovi, kot je DNK leta 1953, se je začela razvijati molekularna biologija. Nadalje v procesu raziskav so znanstveniki ugotovili, kako se DNK rekombinira, njena sestava in kako je strukturiran naš človeški genom.
Vsak dan se na molekularni ravni odvijajo kompleksni procesi. Kako je zgrajena molekula DNK, iz česa je sestavljena? In kakšno vlogo imajo v celici molekule DNK? Pogovorimo se podrobneje o vseh procesih, ki se pojavljajo znotraj dvojne verige.
Kaj je dedna informacija?
Kje se je torej vse začelo? Leta 1868 so ga našli v jedrih bakterij. In leta 1928 je N. Koltsov predstavil teorijo, da so vse genetske informacije o živem organizmu šifrirane v DNK. Nato sta J. Watson in F. Crick leta 1953 našla model danes dobro znane vijačnice DNK, za kar sta zasluženo prejela priznanje in nagrado - Nobelova nagrada.
Kaj sploh je DNK? Ta snov je sestavljena iz dveh združenih niti ali bolje rečeno spiral. Del takšne verige z določenimi informacijami se imenuje gen.

DNK hrani vse informacije o tem, kakšne vrste beljakovin bodo nastale in v kakšnem vrstnem redu. Makromolekula DNK je materialni nosilec neverjetno obsežnih informacij, ki so zapisane v strogem zaporedju posameznih zidakov - nukleotidov. Skupno so 4 nukleotidi, ki se kemično in geometrijsko dopolnjujejo. To načelo dopolnjevanja ali komplementarnosti v znanosti bomo opisali kasneje. To pravilo ima ključno vlogo pri kodiranju in dekodiranju genetskih informacij.
Ker je veriga DNK neverjetno dolga, v tem zaporedju ni nobenih ponovitev. Vsako živo bitje ima svojo edinstveno nit DNK.
Funkcije DNK
Funkcije vključujejo shranjevanje dednih informacij in njihov prenos na potomce. Brez te funkcije se genom vrste ne bi mogel ohraniti in razviti tisoče let.

Organizmi, ki so bili podvrženi resnim genskim mutacijam, bolj verjetno ne bodo preživeli ali bodo izgubili sposobnost ustvarjanja potomcev. Tako pride do naravne zaščite pred propadanjem vrste. Še ena pomembna pomembna funkcija
— izvajanje shranjenih informacij. Celica ne more ustvariti niti enega vitalnega proteina brez teh navodil, ki so shranjena v dvojni verigi.
Sestava nukleinske kisline
- Zdaj je zagotovo znano, iz česa so sestavljeni sami nukleotidi – gradniki DNK. Vsebujejo 3 snovi:
- Ortofosforna kislina.
- Dušikova baza. Pirimidinske baze – ki imajo samo en obroč. Ti vključujejo timin in citozin. Purinske baze, ki vsebujejo 2 obroča. To sta gvanin in adenin.
saharoza. DNA vsebuje deoksiribozo, RNA vsebuje ribozo.
Število nukleotidov je vedno enako številu dušikovih baz. V posebnih laboratorijih nukleotid razgradijo in iz njega izolirajo dušikovo bazo. Tako se preučujejo posamezne lastnosti teh nukleotidov in morebitne mutacije v njih.
Ravni organizacije dednih informacij

Obstajajo 3 ravni organizacije: genetska, kromosomska in genomska. Vse informacije, potrebne za sintezo novega proteina, so vsebovane v majhnem delu verige - genu. To pomeni, da gen velja za najnižjo in najpreprostejšo raven kodiranja informacij.
Geni pa se sestavijo v kromosome. Zahvaljujoč tej organizaciji nosilca dednega materiala se skupine lastnosti izmenjujejo po določenih zakonitostih in prenašajo iz ene generacije v drugo. Vedeti je treba, da je v telesu neverjetno veliko genov, vendar se informacije ne izgubijo niti ob večkratni rekombinaciji.
- Obstaja več vrst genov:
- Glede na njihov funkcionalni namen obstajata 2 vrsti: strukturna in regulativna zaporedja;
Glede na njihov vpliv na procese, ki potekajo v celici, ločijo: supervitalne, letalne, pogojno letalne gene ter mutatorske in antimutatorske gene.
Kaj je genom? To je ime, dano celotnemu nizu nukleotidnih zaporedij v celici organizma. Genom označuje celotno vrsto, ne posameznika.
Kaj je človeška genetska koda?
Dejstvo je, da je ves ogromen potencial človekov razvoj postavljena že v obdobju spočetja. Vse dedne informacije, ki so potrebne za razvoj zigote in rast otroka po rojstvu, so šifrirane v genih. Odseki DNK so najosnovnejši nosilci dednih informacij.
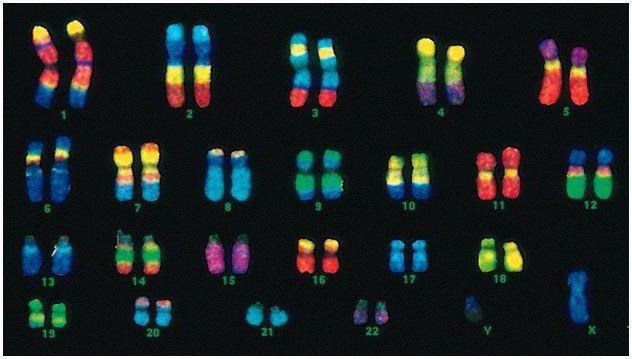
Ljudje imamo 46 kromosomov ali 22 somatskih parov in po en kromosom, ki določa spol, od vsakega starša. V tem diploidnem naboru kromosomov je zakodirana celotna fizična podoba človeka, njegove psihične in fizične sposobnosti ter dovzetnost za bolezni. Somatski kromosomi se navzven ne razlikujejo, vendar nosijo različne informacije, saj je eden od očeta, drugi od matere.
Moška koda se od ženske razlikuje v zadnjem paru kromosomov - XY. Samski diploidni niz je zadnji par, XX. Moški od biološke matere prejmejo en kromosom X, ki ga nato prenesejo na svoje hčere. Spolni Y kromosom prenesel na sinove.
Človeški kromosomi se zelo razlikujejo po velikosti. Na primer, najmanjši par kromosomov je št. 17. In največji par je 1 in 3.
Premer dvojne vijačnice pri človeku je le 2 nm. DNK je zvita tako tesno, da se prilega majhnemu jedru celice, čeprav bi bila dolga do 2 metra, če bi bila odvita. Dolžina vijačnice je več sto milijonov nukleotidov.
Kako se prenaša genetska koda?
Torej, kakšno vlogo imajo molekule DNK pri delitvi celic? Geni - nosilci dednih informacij - se nahajajo v vsaki celici telesa. Da bi svojo kodo prenesli na hčerinski organizem, mnoga bitja razdelijo svojo DNK na 2 enaki vijačnici. To se imenuje replikacija. Med procesom replikacije se DNK odvija in posebni »stroji« dokončajo vsako verigo. Ko se genska vijačnica razcepi, se začnejo deliti jedro in vsi organeli, nato pa celotna celica.
Toda ljudje imamo drugačen proces prenosa genov - spolni. Značilnosti očeta in matere so mešane, nova genetska koda vsebuje podatke obeh staršev.
Shranjevanje in prenašanje dednih informacij je možno zaradi zapletene organizacije vijačnice DNK. Konec koncev, kot smo rekli, je struktura beljakovin šifrirana v genih. Ko je enkrat ustvarjena v času spočetja, se bo ta koda kopirala vse življenje. Kariotip (osebni nabor kromosomov) se med obnavljanjem celic organa ne spreminja. Prenos informacij se izvaja s pomočjo spolnih gamet - moških in žensk.
Samo virusi, ki vsebujejo eno verigo RNA, niso sposobni prenesti svojih informacij na svoje potomce. Zato za razmnoževanje potrebujejo človeške ali živalske celice.
Izvajanje dednih informacij
V jedru celice se nenehno pojavljajo pomembne procese. Vse informacije, zapisane v kromosomih, se uporabljajo za gradnjo beljakovin iz aminokislin. Toda veriga DNK nikoli ne zapusti jedra, zato potrebuje pomoč druge pomembne spojine: RNK. RNK je tista, ki lahko prodre skozi jedrno membrano in sodeluje z verigo DNK.
Skozi interakcijo DNK in 3 tipov RNK se realizirajo vse kodirane informacije. Na kateri ravni poteka implementacija dednih informacij? Vse interakcije potekajo na ravni nukleotidov. Messenger RNA kopira del verige DNA in to kopijo pripelje do ribosoma. Tu se začne sinteza nove molekule iz nukleotidov.
Da bi mRNA kopirala potreben del verige, se vijačnica odvije in nato, po zaključku procesa rekodiranja, ponovno obnovi. Poleg tega lahko ta proces poteka hkrati na dveh straneh 1 kromosoma.
Načelo komplementarnosti
Sestavljeni so iz 4 nukleotidov - adenin (A), gvanin (G), citozin (C), timin (T). Povezani so z vodikovimi vezmi po pravilu komplementarnosti. Delo E. Chargaffa je pomagalo vzpostaviti to pravilo, saj je znanstvenik opazil nekaj vzorcev v obnašanju teh snovi. E. Chargaff je odkril, da je molsko razmerje med adeninom in timinom enako ena. In na enak način je razmerje med gvaninom in citozinom vedno enako ena.
Na podlagi njegovega dela so genetiki oblikovali pravilo za interakcijo nukleotidov. Pravilo komplementarnosti pravi, da se adenin veže samo s timinom, gvanin pa samo s citozinom. Med dekodiranjem vijačnice in sintezo novega proteina v ribosomu to pravilo menjave pomaga hitro najti potrebno aminokislino, ki je pritrjena na prenosno RNA.
RNA in njene vrste
Kaj je dedna informacija? nukleotidov v dvojni verigi DNK. Kaj je RNA? Kaj je njena služba? RNK ali ribonukleinska kislina pomaga pridobivati informacije iz DNK, jih dekodirati in po principu komplementarnosti ustvarjati celicam potrebne beljakovine.
Skupaj obstajajo 3 vrste RNA. Vsak od njih opravlja strogo svojo funkcijo.
- Informacijski (mRNA), ali imenovana tudi matrika. Gre naravnost v središče celice, v jedro. V enem od kromosomov najde potreben genetski material za izgradnjo beljakovine in kopira eno od stranic dvojne verige. Kopiranje spet poteka po principu komplementarnosti.
- Transport je majhna molekula, ki ima na eni strani dekoderje nukleotidov, na drugi strani pa aminokisline, ki ustrezajo osnovni kodi. Naloga tRNA je, da jo dostavi v "delavnico", to je v ribosom, kjer sintetizira potrebno aminokislino.
- rRNA je ribosomska. Nadzoruje količino proizvedenih beljakovin. Sestavljen je iz dveh delov - aminokislinskega in peptidnega dela.
Edina razlika pri dekodiranju je, da RNA nima timina. Namesto timina je tu prisoten uracil. Toda med procesom sinteze beljakovin tRNA še vedno pravilno namesti vse aminokisline. Če pride do napak pri dekodiranju informacij, pride do mutacije.
Popravilo poškodovane molekule DNA
Postopek obnavljanja poškodovane dvojne niti se imenuje popravilo. Med procesom popravljanja se poškodovani geni odstranijo.

Nato se zahtevano zaporedje elementov natančno reproducira in odreže nazaj na isto mesto na verigi, od koder je bilo odstranjeno. Vse to se zgodi zahvaljujoč posebnim kemikalije- encimi.
Zakaj pride do mutacij?
Zakaj začnejo nekateri geni mutirati in prenehajo opravljati svojo nalogo – shranjevanje vitalnih dednih informacij? To se zgodi zaradi napake pri dekodiranju. Na primer, če adenin pomotoma zamenjamo s timinom.
Obstajajo tudi kromosomske in genomske mutacije. Do kromosomskih mutacij pride, ko se deli dednih informacij izgubijo, podvojijo ali celo prenesejo in vstavijo v drug kromosom.

Najresnejše so genomske mutacije. Njihov vzrok je sprememba števila kromosomov. To pomeni, da je namesto para - diploidnega niza v kariotipu prisoten triploidni niz.
Najbolj znan primer triploidne mutacije je Downov sindrom, pri katerem je osebni nabor kromosomov 47. Pri takih otrocih se namesto 21. para oblikujejo 3 kromosomi.
Obstaja tudi znana mutacija, imenovana poliploidija. Toda poliploidija se pojavlja le pri rastlinah.
Molekula DNK je sestavljena iz dveh verig, ki tvorita dvojno vijačnico. Njegovo strukturo sta leta 1953 prva razvozlala Francis Crick in James Watson.
Sprva je molekula DNK, sestavljena iz para nukleotidnih verig, zvitih druga okoli druge, sprožila vprašanja o tem, zakaj ima to posebno obliko. Znanstveniki temu pojavu pravijo komplementarnost, kar pomeni, da se v njegovih verigah lahko nahajajo le določeni nukleotidi drug nasproti drugega. Na primer, adenin je vedno nasproti timinu, gvanin pa vedno nasproti citozinu. Ti nukleotidi molekule DNA se imenujejo komplementarni.
Shematično je prikazano takole:
T - A
C - G
Ti pari tvorijo kemično nukleotidno vez, ki določa vrstni red aminokislin. V prvem primeru je nekoliko šibkejši. Povezava med C in G je močnejša. Nekomplementarni nukleotidi med seboj ne tvorijo parov.
O zgradbi
Struktura molekule DNK je torej posebna. To obliko ima z razlogom: dejstvo je, da je število nukleotidov zelo veliko in je za namestitev dolgih verig potrebno veliko prostora. Prav zaradi tega je za verige značilen spiralni zasuk. Ta pojav imenujemo spiralizacija, omogoča, da se niti skrajšajo za približno pet do šestkrat.
Telo uporablja nekatere tovrstne molekule zelo aktivno, druge redko. Slednje so poleg spiralizacije podvržene tudi tako »kompaktnemu pakiranju«, kot je superspiralizacija. In potem se dolžina molekule DNK zmanjša za 25-30-krat.

Kaj je "embalaža" molekule?
Proces superzvijanja vključuje histonske proteine. Imajo strukturo in videz koluta niti ali palice. Na njih so navite spiralizirane niti, ki se takoj "kompaktno zapakirajo" in zavzamejo malo prostora. Ko se pojavi potreba po uporabi ene ali druge niti, se odvije iz tuljave, na primer histonskega proteina, in vijačnica se odvije v dve vzporedni verigi. Ko je molekula DNK v tem stanju, je mogoče iz nje prebrati potrebne genetske podatke. Vendar obstaja en pogoj. Pridobivanje informacij je možno le, če ima struktura molekule DNA nezapleteno obliko. Kromosomi, ki so dostopni za branje, se imenujejo evhromatini, in če so superzviti, potem so že heterokromatini.
Nukleinske kisline
Nukleinske kisline so tako kot beljakovine biopolimeri. Glavna funkcija je shranjevanje, izvajanje in prenos dednih (genetskih informacij). Obstajajo dve vrsti: DNA in RNA (deoksiribonukleinska in ribonukleinska). Monomeri v njih so nukleotidi, od katerih vsak vsebuje ostanek fosforne kisline, petogljikov sladkor (dezoksiriboza/riboza) in dušikovo bazo. Koda DNK vključuje 4 vrste nukleotidov - adenin (A) / gvanin (G) / citozin (C) / timin (T). Razlikujejo se po dušikovi bazi, ki jo vsebujejo.
V molekuli DNK je lahko število nukleotidov ogromno - od nekaj tisoč do deset in sto milijonov. Takšne velikanske molekule je mogoče pregledati z elektronskim mikroskopom. V tem primeru boste lahko videli dvojno verigo polinukleotidnih verig, ki so med seboj povezane z vodikovimi vezmi dušikovih baz nukleotidov.

Raziskovanje
Med raziskavo so znanstveniki ugotovili, da se vrste molekul DNK v različnih živih organizmih razlikujejo. Ugotovljeno je bilo tudi, da se enoverižni gvanin lahko veže le na citozin, timin pa na adenin. Razporeditev nukleotidov v eni verigi strogo ustreza vzporedni. Zahvaljujoč tej komplementarnosti polinukleotidov je molekula DNA sposobna podvojitve in samoreprodukcije. Toda najprej se komplementarne verige pod vplivom posebnih encimov, ki uničijo seznanjene nukleotide, razhajajo, nato pa se v vsaki od njih začne sinteza manjkajoče verige. To se zgodi zaradi razpoložljivih velike količine prostih nukleotidov v vsaki celici. Posledica tega je, da namesto "matične molekule" nastaneta dve "hčerinski", enaki po sestavi in strukturi, koda DNK pa postane prvotna. Ta proces je predhodnik delitve celic. Zagotavlja prenos vseh dednih podatkov iz matičnih celic v hčerinske celice ter na vse naslednje generacije.
Kako se bere genska koda?
Danes se ne izračuna le masa molekule DNK - mogoče je izvedeti tudi bolj zapletene podatke, ki prej niso bili dostopni znanstvenikom. Preberete lahko na primer informacije o tem, kako organizem uporablja lastno celico. Seveda je ta informacija najprej v kodirani obliki in ima obliko določene matrice, zato jo je treba prenesti na poseben medij, to je RNA. Ribonukleinska kislina lahko prodre v celico skozi jedrsko membrano in prebere kodirane informacije v njej. RNK je torej nosilec skritih podatkov iz jedra v celico, od DNK pa se razlikuje po tem, da namesto deoksiriboze vsebuje ribozo, namesto timina pa uracil. Poleg tega je RNA enoverižna.

sinteza RNA
Poglobljena analiza DNK je pokazala, da RNK, potem ko zapusti jedro, vstopi v citoplazmo, kjer se lahko kot matriks integrira v ribosome (posebne encimske sisteme). Na podlagi prejetih informacij lahko sintetizirajo ustrezno zaporedje beljakovinskih aminokislin. O kakšni vrsti organska spojina mora biti pritrjen na nastajajočo beljakovinsko verigo, se ribosom nauči iz tripletne kode. Vsaka aminokislina ima svoj specifični triplet, ki jo kodira.
Po končani tvorbi verige dobi določeno prostorsko obliko in se spremeni v beljakovino, ki je sposobna opravljati svoje hormonske, konstrukcijske, encimske in druge funkcije. Za vsak organizem je genski produkt. Iz njega se določajo vse vrste lastnosti, lastnosti in manifestacije genov.

Geni
Procesi sekvenciranja so bili razviti predvsem za pridobivanje informacij o tem, koliko genov ima molekula DNK v svoji strukturi. In čeprav so raziskave znanstvenikom omogočile velik napredek v tej zadevi, njihovega natančnega števila še ni mogoče vedeti.
Še pred nekaj leti so domnevali, da molekule DNK vsebujejo približno 100 tisoč genov. Malo kasneje se je številka zmanjšala na 80 tisoč, leta 1998 pa so genetiki izjavili, da je v eni DNK prisotnih le 50 tisoč genov, kar je le 3% celotne dolžine DNK. Toda najnovejši zaključki genetikov so bili osupljivi. Zdaj trdijo, da genom vključuje 25-40 tisoč teh enot. Izkazalo se je, da je le 1,5 % kromosomske DNK odgovorno za kodiranje beljakovin.
Raziskave se tu niso ustavile. Vzporedna ekipa strokovnjakov za genski inženiring je ugotovila, da je število genov v eni molekuli natančno 32 tisoč. Kot lahko vidite, je še vedno nemogoče dobiti dokončen odgovor. Protislovij je preveč. Vsi raziskovalci se zanašajo samo na svoje rezultate.
Je obstajala evolucija?
Kljub dejstvu, da ni dokazov o evoluciji molekule (ker je struktura molekule DNK krhka in majhna), so znanstveniki vseeno podali eno domnevo. Na podlagi laboratorijskih podatkov so izrazili naslednjo različico: molekula na začetni fazi njegovega videza je prevzel obliko preprostega samopodvajajočega se peptida, ki je vseboval do 32 aminokislin, najdenih v starodavnih oceanih.
Po samopodvajanju so molekule zaradi sil naravne selekcije pridobile sposobnost zaščite pred zunanjimi elementi. Začele so živeti dlje in se razmnoževati v večjih količinah. Molekule, ki so se znašle v lipidnem mehurčku, so imele vse možnosti, da se razmnožijo. Kot rezultat niza zaporednih ciklov so lipidni mehurčki dobili obliko celičnih membran, nato pa - vse znani delci. Treba je opozoriti, da je danes vsak odsek molekule DNK zapletena in jasno delujoča struktura, katere vse značilnosti znanstveniki še niso v celoti raziskane.

Sodobni svet
Pred kratkim so znanstveniki iz Izraela razvili računalnik, ki lahko izvede trilijone operacij na sekundo. Danes je to najhitrejši avto na Zemlji. Vsa skrivnost je v tem, da inovativno napravo poganja DNK. Profesorji pravijo, da bodo takšni računalniki v bližnji prihodnosti lahko celo proizvajali energijo.
Pred enim letom so strokovnjaki z Weizmannovega inštituta v Rehovotu (Izrael) napovedali ustvarjanje programabilnega molekularnega računalniškega stroja, sestavljenega iz molekul in encimov. Z njimi so zamenjali silicijeve mikročipe. Do danes je ekipa še napredovala. Sedaj lahko samo ena molekula DNK računalniku zagotovi potrebne podatke in potrebno gorivo.
Biokemični »nanoračunalniki« niso fantazija, ampak že obstajajo v naravi in se manifestirajo v vsakem živem bitju. Toda pogosto jih ne upravljajo ljudje. Človek še ne more delati z genomom katere koli rastline, da bi izračunal, recimo, število "Pi".
Zamisel o uporabi DNK za shranjevanje/obdelavo podatkov je znanstvenikom prvič prišla na misel leta 1994. Takrat je bila molekula uporabljena za rešitev preprostega matematičnega problema. Od takrat so številne raziskovalne skupine predlagale različne projekte, povezane z računalniki DNK. Toda tukaj so vsi poskusi temeljili le na energijski molekuli. Takšnega računalnika ne morete videti s prostim očesom; videti je kot prozorna raztopina vode v epruveti. V njem ni mehanskih delov, ampak le trilijone biomolekularnih naprav – in to le v eni kapljici tekočine!
Človeški DNK
Ljudje so postali pozorni na vrsto človeške DNK leta 1953, ko so znanstveniki lahko svetu prvič predstavili model dvoverižne DNK. Za to sta Kirk in Watson prejela Nobelovo nagrado, saj je to odkritje postalo temeljno v 20. stoletju.
Sčasoma so seveda dokazali, da lahko strukturirana človeška molekula izgleda ne samo kot v predlagani različici. Po podrobnejši analizi DNK so odkrili A-, B- in levosučno obliko Z-. Oblika A- je pogosto izjema, saj nastane le ob pomanjkanju vlage. Toda to je mogoče le v laboratorijskih študijah; za naravno okolje je to nenormalno; v živi celici se tak proces ne more zgoditi.
Oblika B je klasična in je znana kot dvojna desnosučna veriga, oblika Z pa ni le zavita v nasprotni smeri v levo, ampak ima tudi bolj cik-cak videz. Znanstveniki so identificirali tudi obliko G-kvadrupleksa. Njegova struktura nima 2, ampak 4 niti. Po mnenju genetikov se ta oblika pojavlja na območjih, kjer je presežek gvanina.

Umetna DNK
Danes že obstaja umetna DNK, ki je identična kopija prave; popolnoma sledi strukturi naravne dvojne vijačnice. Toda za razliko od neokrnjenega polinukleotida ima umetni samo dva dodatna nukleotida.
Ker je bila sinhronizacija ustvarjena na podlagi informacij, pridobljenih z različnimi študijami prave DNK, se lahko tudi kopira, samopodvaja in razvija. Strokovnjaki se že približno 20 let ukvarjajo z ustvarjanjem takšne umetne molekule. Rezultat je neverjeten izum, ki lahko uporablja genetsko kodo na enak način kot naravni DNK.
Obstoječim štirim dušikovim bazam so genetiki dodali še dve dodatni, ki sta nastali s kemijsko modifikacijo naravnih baz. Za razliko od naravne DNK se je izkazalo, da je umetna DNK precej kratka. Vsebuje le 81 baznih parov. Vendar pa se tudi razmnožuje in razvija.
Replikacija dobljene molekule umetno, poteka zahvaljujoč verižni reakciji s polimerazo, vendar se zaenkrat to ne dogaja samostojno, temveč s posredovanjem znanstvenikov. Omenjeni DNK samostojno dodajajo potrebne encime in jo dajo v posebej pripravljen tekoči medij.
Končni rezultat
Na proces in končni rezultat razvoja DNK lahko vplivajo različni dejavniki, na primer mutacije. Zaradi tega je treba preučiti vzorce snovi, tako da je rezultat analize zanesljiv in zanesljiv. Primer je test očetovstva. Vendar ne moremo kaj, da se ne bi veselili, da so incidenti, kot je mutacija, redki. Kljub temu se vzorci snovi vedno znova preverjajo, da bi na podlagi analize dobili natančnejše podatke.

DNK rastline
Zahvaljujoč tehnologijam visokega sekvenciranja (HTS) je bila revolucija dosežena tudi na področju genomike – mogoča je tudi ekstrakcija DNK iz rastlin. Seveda pridobivanje molekulske mase DNK iz rastlinskega materiala visoke kakovosti povzroča nekaj težav zaradi veliko število kopije mitohondrijske in kloroplastne DNA, kot tudi visoki ravni polisaharidi in fenolne spojine. Za izolacijo strukture, ki jo obravnavamo v tem primeru, se uporabljajo različne metode.
Vodikova vez v DNK
Vodikova vez v molekuli DNK je odgovorna za elektromagnetno privlačnost, ki nastane med pozitivno nabitim vodikovim atomom, ki je vezan na elektronegativni atom. Ta dipolna interakcija ne izpolnjuje kriterija kemična vez. Lahko pa se pojavi intermolekularno ali v različnih delih molekule, tj. intramolekularno.

Atom vodika se veže na elektronegativni atom, ki je darovalec vezi. Elektronegativni atom je lahko dušik, fluor ali kisik. On – z decentralizacijo – pritegne k sebi elektronski oblak iz vodikovega jedra in naredi vodikov atom (delno) pozitivno nabit. Ker je velikost H majhna v primerjavi z drugimi molekulami in atomi, je tudi naboj majhen.
dekodiranje DNK
Pred dešifriranjem molekule DNK znanstveniki najprej vzamejo ogromno celic. Za najbolj natančno in uspešno delo približno milijon jih je potrebnih. Rezultati, pridobljeni med študijo, se nenehno primerjajo in beležijo. Danes dekodiranje genoma ni več redkost, temveč dostopen postopek.

Seveda je dešifriranje genoma ene same celice nepraktična naloga. Podatki, pridobljeni med takimi študijami, znanstvenikov ne zanimajo. Vendar je pomembno razumeti, da vse trenutno obstoječe metode dekodiranja kljub svoji kompleksnosti niso dovolj učinkovite. Omogočajo le branje 40-70 % DNK.
Vendar pa so profesorji s Harvarda nedavno objavili metodo, s katero je mogoče dešifrirati 90 % genoma. Tehnika temelji na dodajanju začetnih molekul izoliranim celicam, s pomočjo katerih se začne replikacija DNA. Toda tudi te metode ni mogoče šteti za uspešno; treba jo je še izpopolniti, preden jo lahko odkrito uporabimo v znanosti.
DNA (deoksiribonukleinska kislina) se nanaša (skupaj z RNA), ki so polimeri, ali natančneje polinukleotidi (monomer - nukleotid).
DNK je odgovorna za shranjevanje in prenos genetske kode med celično delitvijo. Preko molekul DNK se uresničujeta dednost in variabilnost. Vse vrste RNA se sintetizirajo iz DNA. Poleg tega različne vrste RNA skupaj zagotavljajo sintezo celičnih proteinov, tj. izvajajo genetske informacije.
V evkariontskih celicah se velika večina DNK nahaja v jedru, kjer se kompleksira s posebnimi proteini in tvori kromosome. V prokariontskih celicah je ena velika krožna (ali linearna) molekula DNA (tudi v kompleksu z beljakovinami). Poleg tega imajo evkariontske celice lastno DNK v mitohondrijih in kloroplastih.
V primeru DNK je vsak nukleotid sestavljen iz 1) dušikove baze, ki je lahko adenin, gvanin, citozin ali timin, 2) deoksiriboze, 3) fosforne kisline.
Zaporedje nukleotidov v verigi DNA določa primarno strukturo molekule. Za DNK je značilna sekundarna struktura molekule v obliki dvojne vijačnice (najpogosteje desnosučne). V tem primeru sta dve verigi DNK med seboj povezani z vodikovimi vezmi, ki nastanejo med komplementarnimi dušikovimi bazami.
Adenin je komplementaren timinu, gvanin pa citozinu. Med adeninom in timinom se tvorita dve vodikovi vezi, med gvaninom in citozinom pa tri. Tako sta gvanin in citozin med seboj nekoliko tesneje povezana (čeprav so vodikove vezi načeloma šibke). Število vezi je določeno s strukturnimi značilnostmi molekul.
Adenin in gvanin sta purina in sta sestavljena iz dveh obročev. Timin in citozin sta pirimidinski bazi z enim obročem. Tako so med hrbtenicama (sestavljenimi iz izmenjujočih se deoksiriboze in fosforne kisline) dveh verig DNA za kateri koli par nukleotidov različnih verig vedno trije obroči (ker je purin z dvema obročema vedno komplementaren le določenemu enojnemu obroču pirimidin). To omogoča, da je širina med verigami molekule DNK ves čas enaka (približno 2,3 nm).
V enem obratu vijačnice je približno 10 nukleotidov. Dolžina enega nukleotida je približno 0,34 nm. Dolžina molekul DNK je običajno ogromna in presega milijone nukleotidov. Zato je DNK podvržena različnim stopnjam »superzvijanja«, da bi se bolj kompaktno prilegala celičnemu jedru.
Pri branju informacij iz DNK (to je sintetiziranju RNK na njej se ta proces imenuje prepisovanje) pride do despiralizacije (obraten proces spiralizacije), dve verigi se razhajata pod delovanjem posebnega encima. Vodikove vezi so šibke, zato pride do ločevanja in poznejšega zamreženja verig z malo porabe energije. RNA se sintetizira iz DNA po enakem principu komplementarnosti. Le namesto timina v RNA je adeninu komplementaren uracil.
Genetska koda zapisan na molekulah DNK, sestoji iz trojčkov (zaporedja treh nukleotidov), ki predstavljajo eno aminokislino (proteinski monomer). Vendar pa večina DNK ne kodira beljakovin. Pomen takih delov molekule je različen in večinoma ni popolnoma razumljen.
Preden se celica deli, se količina DNK vedno podvoji. Ta proces se imenuje podvajanje. Je polkonzervativne narave: verige ene molekule DNK se razhajajo in vsaka se dopolni s svojo novo komplementarno verigo. Kot rezultat, iz ene dvoverižne molekule DNA dobimo dve dvoverižni DNA, ki sta enaki prvi.
V DNK so polinukleotidne verige večsmerne, tj. kjer ima ena veriga 5" konec (ostanek fosforne kisline je pritrjen na peti ogljikov atom deoksiriboze), druga ima 3" konec (ogljik brez fosforne kisline).
Med replikacijo in transkripcijo poteka sinteza vedno v smeri od 5" konca do 3" konca, saj se novi nukleotidi lahko pritrdijo samo na prosti 3" ogljikov atom.
Struktura in vloga DNK kot snovi, ki je odgovorna za dedno informacijo, je bila pojasnjena v 40-ih in 50-ih letih 20. stoletja. Leta 1953 sta D. Watson in F. Crick določila dvoverižno strukturo DNA. Prej je E. Chargaff ugotovil, da v DNK količina timina vedno ustreza adeninu, količina gvanina pa citozinu.
Sorodni članki
-
Vojaška naselja Puškin okoli Arakcheeva
Aleksej Andrejevič Arakčejev (1769-1834) - ruski državnik in vojskovodja, grof (1799), artilerijski general (1807). Izhajal je iz plemiške družine Arakčejevih. Uveljavil se je pod Pavlom I. in prispeval k njegovi vojaški ...
-
Preprosti fizikalni poskusi doma
Lahko se uporablja pri pouku fizike na stopnjah postavljanja ciljev in ciljev lekcije, ustvarjanja problemskih situacij pri preučevanju nove teme, uporabe novega znanja pri utrjevanju. Predstavitev Zabavni poskusi lahko učenci uporabljajo za...
-
Dinamična sinteza odmičnih mehanizmov Primer sinusnega zakona gibanja odmičnih mehanizmov
Odmični mehanizem je mehanizem z višjim kinematičnim parom, ki ima možnost zagotoviti obstojnost izhodnega člena, struktura pa vsebuje vsaj en člen z delovno površino spremenljive ukrivljenosti. Cam mehanizmi ...
-
Vojna se še ni začela Vse Podkast oddaje Glagolev FM
Predstava Semjona Aleksandrovskega po drami Mihaila Durnenkova "Vojna se še ni začela" je bila uprizorjena v gledališču Praktika. Poroča Alla Shenderova. V zadnjih dveh tednih je to že druga moskovska premiera po besedilu Mihaila Durnenkova....
-
Predstavitev na temo "metodološka soba v dhowu"
| Dekoracija pisarn v predšolski vzgojni ustanovi Zagovor projekta "Novoletna dekoracija pisarne" za mednarodno leto gledališča Bilo je januarja A. Barto Gledališče senc Rekviziti: 1. Velik zaslon (list na kovinski palici) 2. Svetilka za vizažisti...
-
Datumi Olgine vladavine v Rusiji
Po umoru kneza Igorja so se Drevljani odločili, da je odslej njihovo pleme svobodno in da jim ni treba plačevati davka Kijevski Rusiji. Še več, njihov princ Mal se je poskušal poročiti z Olgo. Tako se je želel polastiti kijevskega prestola in sam...