ДНК молекула. Структура на молекулата на ДНК. Наследствена информация: съхранение и предаване. Генетичен код. ДНК верига, където се намират молекули ДНК
Нуклеиновите киселини са високомолекулни вещества, състоящи се от мононуклеотиди, които са свързани помежду си в полимерна верига, използвайки 3 ", 5" - фосфодиестерни връзки и са опаковани в клетки по определен начин.
Нуклеиновите киселини са биополимери от два вида: рибонуклеинова киселина (РНК) и дезоксирибонуклеинова киселина (ДНК). Всеки биополимер се състои от нуклеотиди, които се различават по въглехидратния остатък (рибоза, дезоксирибоза) и една от азотните основи (урацил, тимин). Според тези различия нуклеиновите киселини са получили името си.
Структура на дезоксирибонуклеинова киселина
Нуклеиновите киселини имат първична, вторична и третична структура.
Първична структура на ДНК
Първичната структура на ДНК се нарича линейна полинуклеотидна верига, в която мононуклеотидите са свързани чрез 3 ", 5" -фосфодиестерни връзки. Изходен материал при сглобяване на веригата нуклеинова киселинав клетката се намира нуклеозид 5 "-трифосфат, който в резултат на отстраняването на β и γ остатъци от фосфорна киселина е в състояние да прикрепи 3" въглеродния атом на друг нуклеозид. По този начин 3 "въглеродният атом на една дезоксирибоза ковалентно се свързва с 5" въглероден атом на другата дезоксирибоза чрез един остатък от фосфорна киселина и образува линейна полинуклеотидна верига нуклеинова киселина. Оттук и името: 3 ", 5" -фосфодиестерни връзки. Азотните основи не участват в комбинацията от нуклеотиди от една верига (фиг. 1.).
Такава връзка, между остатъка от молекулата на фосфорната киселина на един нуклеотид и въглехидратите на друг, води до образуването на пентозо-фосфатен скелет на полинуклеотидната молекула, върху който азотните основи са прикрепени една по една отстрани. Тяхната последователност от подреждане във веригите на молекули нуклеинова киселина е строго специфична за клетки от различни организми, т.е. е със специфичен характер (правилото на Чаргаф).
Линейна ДНК верига, чиято дължина зависи от броя на нуклеотидите, включени във веригата, има два края: единият се нарича 3 "край и съдържа свободен хидроксил, а другият, 5" край, съдържа фосфорна киселина остатък. Веригата е поляризирана и може да има посока 5 "-> 3" и 3 "-> 5". Изключение е кръговата ДНК.
Генетичният "текст" на ДНК е съставен от кодови "думи" - тройки нуклеотиди, наречени кодони. Областите на ДНК, които съдържат информация за първичната структура на всички видове РНК, се наричат структурни гени.
Полинуклеодитовите ДНК вериги достигат гигантски размери, така че те са опаковани по определен начин в клетката.
Изучавайки състава на ДНК, Чаргаф (1949) установява важни закони относно съдържанието на отделни ДНК бази. Те помогнаха да се разкрие вторичната структура на ДНК. Тези модели се наричат правила на Чаргаф. Правилата на Чаргаф
Тези правила показват, че при изграждането на ДНК не трябва да се наблюдава доста строго съответствие (сдвояване) между пуринови и пиримидинови основи като цяло, а по -специално тимин с аденин и цитозин с гуанин. Въз основа на тези правила, включително през 1953 г. Уотсън и Крик предлагат модел на вторичната структура на ДНК, наречен двойна спирала (фиг.). |

Вторична структура на ДНК
Вторичната структура на ДНК е двойна спирала, чийто модел е предложен от Д. Уотсън и Ф. Крик през 1953 г.
Предпоставки за създаване на ДНК модел
Първоначалните анализи създадоха впечатлението, че ДНК от произволен произход съдържа и четирите нуклеотида в равни моларни количества. Въпреки това през 40 -те години на миналия век Е. Чаргаф и неговите сътрудници, в резултат на анализа на ДНК, изолирана от различни организми, ясно показаха, че азотните основи се съдържат в тях в различни количествени съотношения. Чаргаф установи, че въпреки че тези съотношения са еднакви за ДНК от всички клетки от един и същ тип организъм, ДНК от различни видове може да се различава значително в съдържанието на определени нуклеотиди. Това предполага, че разликите в съотношението на азотни основи могат да бъдат свързани с някакъв биологичен код. Въпреки че съотношението на отделните пуринови и пиримидинови основи в различните ДНК проби се оказа различно, при сравняване на резултатите от анализа се появи определен модел: във всички проби общото количество пурини беше равно на общото количество пиримидини (А + G = T + C), количеството аденин е равно на количеството тимин (A = T), а количеството на гуанин - количеството цитозин (G = C). ДНК, изолирана от клетки на бозайници, като цяло е по -богата на аденин и тимин и относително по -бедна на гуанин и цитозин, докато при бактериите ДНК е по -богата на гуанин и цитозин и относително по -бедна на аденин и тимин. Тези данни съставляват важна част от фактическия материал, въз основа на който по-късно е изграден модел на структурата на ДНК на Уотсън-Крик.
Друга важна непряка индикация за възможната структура на ДНК са данните на Л. Полинг за структурата на протеиновите молекули. Полинг показа, че са възможни няколко различни стабилни конфигурации на аминокиселинната верига в протеинова молекула. Една от често срещаните конфигурации на пептидната верига, α-спиралата, е правилна спирална структура. При такава структура е възможно образуването на водородни връзки между аминокиселини, разположени на съседни завои на веригата. Полинг описва α-спиралната конфигурация на полипептидната верига през 1950 г. и предполага, че молекулите на ДНК също вероятно имат спирална структура, фиксирана от водородни връзки.
Най-ценната информация за структурата на молекулата на ДНК обаче беше предоставена от резултатите от рентгеновия структурен анализ. Рентгеновите лъчи, преминаващи през кристала на ДНК, претърпяват дифракция, тоест се отклоняват в определени посоки. Степента и естеството на отклонението на лъчите зависят от структурата на самите молекули. Рентгеновата дифракционна картина (фиг. 3) дава на опитното око редица косвени индикации относно структурата на молекулите на изследваното вещество. Анализът на рентгеновите дифракционни модели на ДНК доведе до заключението, че азотните основи (с плоска форма) са подредени като купчина плочи. Рентгеновите дифракционни модели разкриват три основни периода в структурата на кристалната ДНК: 0,34, 2 и 3,4 nm.

ДНК модел на Уотсън-Крик
Въз основа на аналитичните данни на Чаргаф, рентгеновите дифракционни модели, получени от Уилкинс и изследванията на химици, които предоставиха информация за точните разстояния между атомите в молекулата, за ъглите между връзките на даден атом и размера на атоми, Уотсън и Крик започнаха да изграждат физически модели на отделните съставни части на молекулата на ДНК в определен мащаб и да ги „напасват“ помежду си по такъв начин, че получената система да отговаря на различни експериментални данни [показване] .
Още по-рано беше известно, че съседните нуклеотиди в ДНК веригата са свързани чрез фосфодиестерни мостове, свързващи 5'-въглеродния атом на дезоксирибоза на един нуклеотид с 3'-въглеродния атом на дезоксирибоза на следващия нуклеотид. Уотсън и Крик не се съмняват, че периодът от 0,34 nm съответства на разстоянието между последователни нуклеотиди в ДНК веригата. Освен това може да се предположи, че периодът от 2 nm съответства на дебелината на веригата. И за да обяснят коя реална структура отговаря на периода от 3.4 nm, Уотсън и Крик, точно както Полинг по -рано, предложиха веригата да бъде усукана под формата на спирала (или по -точно образува спирална линия, тъй като спирала в строгия смисъл на това думата се получава, когато завоите образуват конусна, а не цилиндрична повърхност в пространството). Тогава периодът от 3,4 nm ще съответства на разстоянието между последователните завои на тази спирала. Такава спирала може да бъде много плътна или донякъде опъната, тоест завоите й могат да бъдат нежни или стръмни. Тъй като периодът от 3,4 nm е точно 10 пъти разстоянието между последователните нуклеотиди (0,34 nm), е ясно, че всеки пълен оборот на спиралата съдържа 10 нуклеотида. От тези данни Уотсън и Крик успяха да изчислят плътността на полинуклеотидна верига, усукана в спирала с диаметър 2 nm, с разстояние между завоите, равно на 3,4 nm. Оказа се, че плътността на такава верига ще бъде половината от действителната плътност на ДНК, която вече беше известна. Трябваше да предположа, че молекулата на ДНК се състои от две нишки - че е двойна спирала от нуклеотиди.
Следващата задача беше, разбира се, да се изясни пространствената връзка между двете вериги, образуващи двойна спирала. След тестване на редица схеми на вериги на техния физически модел, Уотсън и Крик установиха, че всички налични данни най -добре отговарят на вариант, при който две полинуклеотидни спирали се движат в противоположни посоки; в този случай вериги, състоящи се от захарни и фосфатни остатъци, образуват повърхността на двойна спирала, а вътре са разположени пурини и пиримидини. Базите, разположени една срещу друга, принадлежащи към две вериги, са свързани по двойки чрез водородни връзки; именно тези водородни връзки държат веригите заедно, като по този начин фиксират цялостната конфигурация на молекулата.
Двойната спирала на ДНК може да бъде представена като въжена стълба във формата на спирала, така че стъпалата й да останат в хоризонтално положение. Тогава две надлъжни въжета ще съответстват на вериги от захарни и фосфатни остатъци, а напречните ще отговарят на двойки азотни основи, свързани чрез водородни връзки.
В резултат на по -нататъшно проучване на възможните модели, Уотсън и Крик стигнаха до заключението, че всяка "лента" трябва да се състои от един пурин и един пиримидин; с период от 2 nm (което съответства на диаметъра на двойната спирала), няма да има достатъчно място за двата пурина, а двата пиримидина няма да са достатъчно близо един до друг, за да образуват подходящи водородни връзки. Задълбочено проучване на подробен модел показа, че аденинът и цитозинът, съставляващи подходяща по размер комбинация, все още не могат да бъдат локализирани по такъв начин, че между тях да се образуват водородни връзки. Подобни доклади налагат изключването на комбинацията гуанин-тимин, докато комбинациите аденин-тимин и гуанин-цитозин са напълно приемливи. Природата на водородните връзки е такава, че аденинът образува двойка с тимин, а гуанин с цитозин. Тази концепция за специфично сдвояване на база позволи да се обясни "правилото на Чаргаф", според което във всяка молекула на ДНК количеството на аденина винаги е равно на съдържанието на тимин, а количеството на гуанин е равно на количеството на цитозин. Между аденин и тимин се образуват две водородни връзки, а три между гуанин и цитозин. Поради тази специфичност при образуването на водородни връзки срещу всеки аденин в едната верига, тиминът се намира в другата; по същия начин, срещу всеки гуанин може да се намери само цитозин. По този начин веригите се допълват взаимно, тоест последователността на нуклеотидите в едната верига еднозначно определя тяхната последователност в другата. Двете вериги се движат в противоположни посоки и техните крайни фосфатни групи са в противоположните краища на двойната спирала.
В резултат на своите изследвания през 1953 г. Уотсън и Крик предлагат модел на структурата на молекулата на ДНК (фиг. 3), който остава актуален и до днес. Според модела ДНК молекула се състои от две комплементарни полинуклеотидни вериги. Всяка ДНК верига е полинуклеотид, състоящ се от няколко десетки хиляди нуклеотиди. В него съседни нуклеотиди образуват правилен пентозо-фосфатен гръбнак поради свързването на остатъка от фосфорна киселина и дезоксирибоза със силна ковалентна връзка. В този случай азотните основи на една полинуклеотидна верига са подредени в строго определен ред спрямо азотните бази на другата. Редуването на азотни основи в полинуклеотидната верига е нередовно.
Разположението на азотните бази в ДНК веригата е допълващо (от гръцки „комплемент“ - добавяне), т.е. срещу аденин (А) винаги има тимин (Т), а срещу гуанин (G) - само цитозин (С). Това се дължи на факта, че A и T, както и G и C строго съответстват един на друг, т.е. взаимно се допълват. Дава се такава кореспонденция химическа структурабази, позволяващи образуването на водородни връзки в двойка пурин и пиримидин. Има две връзки между A и T, три между G и C. Тези връзки осигуряват частично стабилизиране на молекулата на ДНК в космоса. В този случай стабилността на двойната спирала е правопропорционална на броя на G≡C връзките, които са по -стабилни в сравнение с A = T връзките.
Известната последователност на подреждането на нуклеотиди в една ДНК верига позволява, съгласно принципа на комплементарност, да се установят нуклеотидите на другата верига.
Освен това е установено, че азотни основи с ароматна структура в воден разтворса разположени една над друга, образувайки така или иначе купчина монети. Този процес на образуване на купчини органични молекули се нарича подреждане. Полинуклеотидните вериги на молекулата на ДНК от разглеждания модел Уотсън-Крик имат подобно физико-химично състояние, азотните им основи са разположени под формата на купчина монети, между равнините на които възникват взаимодействия на ван дер Ваалс (взаимодействия на подреждане).
Водородни връзкимежду комплементарни бази (хоризонтално) и взаимодействие на стекиране между равнините на основите в полинуклеотидната верига поради силите на ван дер Ваалс (вертикално) осигурява молекулата на ДНК с допълнителна стабилизация в пространството.
Захарно-фосфатните гръбнаци на двете вериги са обърнати навън, а основите навътре, един към друг. Посоката на веригите в ДНК е антипаралелна (едната от тях има посока 5 "-> 3", другата-3 "-> 5", т.е. 3 "краят на едната верига е срещу 5" края на другата .). Веригите образуват десни спирали с обща ос. Едно завъртане на спиралата е 10 нуклеотида, размерът на намотката е 3,4 nm, височината на всеки нуклеотид е 0,34 nm, а диаметърът на спиралата е 2,0 nm. В резултат на въртенето на една нишка около другата се образуват голям жлеб (с диаметър около 20 Å) и малък жлеб (около 12 Å) на двойната спирала на ДНК. Тази форма на двойната спирала на Уотсън-Крик по-късно се нарича В-форма. В клетките ДНК обикновено съществува във В-формата, която е най-стабилната форма.
ДНК функции
Предложеният модел обяснява много биологични свойствадезоксирибонуклеинова киселина, включително съхранението на генетична информация и разнообразието от гени, осигурено от голямо разнообразие от последователни комбинации от 4 нуклеотида и фактът на съществуването на генетичния код, способността за самовъзпроизвеждане и предаване на генетична информация, осигурена чрез процеса на репликация и внедряването на генетична информация под формата на протеини и всякакви други съединения, образувани от ензимни протеини.
Основни функции на ДНК.
- ДНК е носител на генетична информация, което се гарантира от факта на съществуването на генетичния код.
- Възпроизвеждане и предаване на генетична информация в поколения клетки и организми. Тази функционалност се осигурява от процеса на репликация.
- Реализиране на генетична информация под формата на протеини, както и всякакви други съединения, образувани с помощта на ензимни протеини. Тази функция се осигурява от процесите на транскрипция и превод.

Форми на организация на двуверижна ДНК
ДНК може да образува няколко вида двойни спирали (фиг. 4). В момента вече са известни шест форми (от А до Е и Z-форма).
Структурните форми на ДНК, както установи Розалинд Франклин, зависят от насищането на молекулата на нуклеиновата киселина с вода. При проучвания на ДНК влакна, използващи рентгенов структурен анализ, беше показано, че рентгеновата дифракционна картина коренно зависи от това при каква относителна влажност, при каква степен на насищане с вода на това влакно, се провежда експериментът. Ако влакното е достатъчно наситено с вода, тогава се получава една рентгенова снимка. При изсушаване се появи напълно различен рентгенов модел, който беше много различен от рентгеновия модел на влакното с висока влажност.
ДНК молекулата с висока влажност се нарича В-форма... При физиологични условия (ниска концентрация на сол, висока степен на хидратация), доминиращият структурен тип ДНК е В-формата (основната форма на двуверижна ДНК е моделът на Уотсън-Крик). Стъпката на спиралата на такава молекула е 3,4 nm. На ход има 10 допълващи се двойки под формата на усукани купчини „монети“ - азотни основи. Купчините се държат чрез водородни връзки между две противоположни „монети“ от купчината и са „увити“ в две ленти от скелет на фосфодиестер, усукани в дясна спирала. Плоскостите на азотни основи са перпендикулярни на оста на спиралата. Съседните допълващи се двойки се завъртат един спрямо друг на 36 °. Диаметърът на спиралата е 20 Å, като пуриновият нуклеотид заема 12 Å, а пиримидиновият нуклеотид - 8 Å.
ДНК молекулата с по-ниска влажност се нарича А-форма... А-формата се образува при условия на по-малко висока хидратация и с по-високо съдържание на Na + или K + йони. Тази по-широка конформация с дясна ръка има 11 базови двойки на ход. Плоскостите на азотни основи имат по -силен наклон към оста на спиралата; те се отклоняват от нормалата към оста на спиралата с 20 °. Следователно следва наличието на вътрешна кухина с диаметър 5 Å. Разстоянието между съседните нуклеотиди е 0,23 nm, дължината на намотката е 2,5 nm, а диаметърът на спиралата е 2,3 nm.
Първоначално А-формата на ДНК се смяташе за по-малко важна. По-късно обаче стана ясно, че А-формата на ДНК, както и В-формата, имат огромно биологично значение. РНК-ДНК спирала в комплекса матрица-праймер има А-форма, както и РНК-РНК спирала и РНК структури за фиби (2'-хидроксилната група на рибозата не позволява на молекулите на РНК да образуват В-форма) . А-формата на ДНК е открита в противоречие. Установено е, че А-формата на ДНК е 10 пъти по-устойчива на UV лъчи от В-формата.
А-формата и В-формата се наричат канонични форми на ДНК.
Форми C-Eсъщо с дясна ръка, тяхното образуване може да се наблюдава само в специални експерименти и очевидно те не съществуват in vivo. С-формата на ДНК има структура, подобна на В-ДНК. Броят базови двойки на ход е 9,33, дължината на спиралата е 3,1 nm. Основните двойки са наклонени под ъгъл от 8 градуса спрямо перпендикулярното положение към оста. Вдлъбнатините са сходни по размер с каналите на В-ДНК. В този случай основният жлеб е малко по -плитък, а второстепенният е по -дълбок. Естествените и синтетичните ДНК полинуклеотиди могат да преминат в С-формата.
| Таблица 1. Характеристики на някои видове ДНК структури | |||
| Спирален тип | А | Б | Z |
| Спирална стъпка | 0,32 nm | 3,38 nm | 4.46 nm |
| Спирално усукване | Точно така | Точно така | Наляво |
| Базови двойки на ход | 11 | 10 | 12 |
| Разстояние между базовите равнини | 0,256 nm | 0,338 nm | 0,371 nm |
| Конформация на гликозидна връзка | анти | анти | анти-С syn-g |
| Конформация на фуранозен пръстен | C3 "-endo | C2 "-endo | C3 "-endo-G C2 "-endo-c |
| Ширина на жлеба малък / голям | 1,11 / 0,22 nm | 0,57 / 1,17 nm | 0,2 / 0,88 nm |
| Дълбочина на канала, малка / голяма | 0,26 / 1,30 nm | 0,82 / 0,85 nm | 1,38 / 0,37 nm |
| Диаметър на спиралата | 2,3 nm | 2,0 nm | 1,8 nm |
Структурни елементи на ДНК
(неканонични ДНК структури)
Структурните елементи на ДНК включват необичайни структури, ограничени от някои специални последователности:
|

Z-образна ДНКе открит през 1979 г. при изучаване на хексануклеотид d (CG) 3 -. Открит е от професора в Масачузетския технологичен институт Александър Рич и неговите колеги. Z-формата се превърна в един от най-важните структурни елементи на ДНК поради факта, че нейното образуване се наблюдава в области на ДНК, където пурините се редуват с пиримидини (например 5'-HCGCH-3 '), или в повторения от 5' -СНСН-3 ', съдържащ метилиран цитозин. Съществено условие за образуването и стабилизирането на Z-ДНК е наличието на пуринови нуклеотиди в син-конформацията, редуващи се с пиримидинови бази в антиконформацията.
Естествените молекули на ДНК съществуват главно в дясната В-форма, ако не съдържат последователности от типа (СН) n. Въпреки това, ако такива последователности са част от ДНК, тогава тези области, когато йонната сила на разтвора или катионите, неутрализиращи отрицателния заряд на фосфодиестерния гръбнак, се променят, могат да се трансформират в Z-форма, докато други ДНК области във веригата остават в класическата В-форма. Възможността за такъв преход показва, че двете вериги в двойната спирала на ДНК са в динамично състояние и могат да се развиват една спрямо друга, преминавайки от дясната форма наляво и обратно. Биологичните последици от такава лабилност, която позволява конформационни трансформации на структурата на ДНК, все още не са напълно изяснени. Смята се, че Z-ДНК регионите играят роля в регулирането на експресията на някои гени и участват в генетичната рекомбинация.
Z-формата на ДНК е лява двойна спирала, в която гръбнакът на фосфодиестера е разположен по зигзаг по оста на молекулата. Оттук и името на молекулата (зигзаг) -ДНК. Z-ДНК е най-малко усуканата (12 двойки основи на оборот) и най-тънката позната в природата. Разстоянието между съседни нуклеотиди е 0,38 nm, дължината на намотката е 4,56 nm, а диаметърът на Z-ДНК е 1,8 nm. В допълнение, външният вид на тази молекула на ДНК се отличава с наличието на единичен жлеб.
Z-форма ДНК е открита в прокариотни и еукариотни клетки. В момента са получени антитела, които могат да различат Z-формата от В-формата на ДНК. Тези антитела се свързват със специфични области на гигантските хромозоми на клетките на слюнчените жлези на Drosophila (Dr. melanogaster). Реакцията на свързване е лесна за проследяване поради необичайната структура на тези хромозоми, при които по -плътните области (дискове) контрастират с по -малко плътните области (междудискове). Секциите на Z-ДНК са разположени в междулентите. От това следва, че Z-формата действително съществува в естествени условия, въпреки че размерите на отделните участъци на Z-формата все още са неизвестни.

(превключватели) са най-известните и често срещани базови последователности в ДНК. Палиндромът е дума или фраза, която се чете отляво надясно и обратно по същия начин. Примери за такива думи или фрази са: SHALASH, KAZAK, POTOP и ROSE FALLED НА AZOR'S Law. Когато се прилага към региони на ДНК, този термин (палиндром) означава същото редуване на нуклеотиди по веригата отдясно наляво и отляво надясно (като буквите в думата „хижа“ и т.н.).
Палиндромът се характеризира с наличието на обърнати повторения на базови последователности със симетрия от втори ред по отношение на две ДНК вериги. Такива последователности, по напълно разбираема причина, се допълват самостоятелно и са склонни да образуват фиби или кръстообразни структури (фиг.). Фибите помагат на регулаторните протеини да разпознаят мястото, където генетичният текст на хромозомната ДНК се отписва.
В случаите, когато инвертирано повторение присъства на една и съща верига на ДНК, тази последователност се нарича огледално повторение. Огледалните повторения не притежават свойствата на самодопълване и следователно не са способни да образуват фиби или кръстообразни структури. Последователности от този тип се намират на практика във всички големи молекули на ДНК и могат да варират от само няколко двойки основи до няколко хиляди бази двойки.
Наличието на палиндроми под формата на кръстовидни структури в еукариотни клетки не е доказано, въпреки че редица кръстовидни структури са открити in vivo в клетки на Е. coli. Наличието на самостоятелно комплементарни последователности в РНК или едноверижна ДНК е основната причина за сгъването на нуклеинова верига в разтвори в определена пространствена структура, характеризираща се с образуването на много „фиби“.

Н-образна ДНКе спирала, образувана от три нишки на ДНК - тройната спирала на ДНК. Това е комплекс от двойната спирала на Уотсън-Крик с третата едноверижна ДНК верига, която се вписва в нейния голям жлеб, с образуването на така наречената двойка Хогстийн.
Образуването на такъв триплекс става в резултат на сгъването на двойната спирала на ДНК по такъв начин, че половината от нейното сечение остава под формата на двойна спирала, а другата половина е изключена. В този случай една от изключените спирали образува нова структура с първата половина на двойната спирала - тройна спирала, а втората се оказва неструктурирана, под формата на едноверижна секция. Характерна особеност на този структурен преход е острата зависимост от рН на средата, чиито протони стабилизират новата структура. Поради тази функция нова структураполучи името на Н-формата на ДНК, чието образуване е открито в свръхсвързани плазмиди, съдържащи хомопурин-хомопиримидинови области, които са огледално повторение.
В по-нататъшни проучвания, възможността за структурен преход на някои хомопурин-хомопиримидинови двуверижни полинуклеотиди с образуване на триверижна структура, съдържаща:
- една хомопуринова и две хомопиримидинови нишки ( Py-Pu-Py триплекс) [Взаимодействие на Hoogsteen].
Съставните блокове на триплекса Py-Pu-Py са каноничните триади, изоморфни на CGC + и TAT. Стабилизирането на триплекса изисква протониране на триадата CGC +; следователно тези триплекси зависят от рН на разтвора.
- една хомопиримидинова и две хомопуринови нишки ( Py-Pu-Pu триплекс) [обратно взаимодействие на Hoogsteen].
Съставните блокове на триплекса Py-Pu-Pu са канонично изоморфни на триадите CGG и TAA. Съществено свойство на триплексите Py-Pu-Pu е зависимостта на тяхната стабилност от наличието на двойно заредени йони и са необходими различни йони за стабилизиране на триплекси от различни последователности. Тъй като образуването на Py-Pu-Pu триплекси не изисква протониране на съставните им нуклеотиди, такива триплекси могат да съществуват при неутрално рН.
Забележка: директното и обратното взаимодействие на Hoogsteen се обяснява със симетрията на 1-метилтимин: въртенето на 180 ° води до факта, че атомът O4 е заменен с атома O2, докато системата от водородни връзки е запазена.
Има два вида тройни спирали:
- успоредни тройни спирали, при които полярността на третата нишка съвпада с полярността на хомопуриновата верига на дуплекса Уотсън-Крик
- антипаралелни тройни спирали, при които полярностите на третата и хомопуриновите вериги са противоположни.

G-четворка- 4-спирална ДНК. Такава структура се формира, ако има четири гуанина, които образуват така наречения G-квадруплекс-кръгъл танц от четири гуанина.
Първите намеци за възможността за формиране на такива структури са получени много преди пробивната работа на Уотсън и Крик - още през 1910 г. Тогава немският химик Ивар Банг открил, че един от компонентите на ДНК - гуанозовата киселина - образува гелове при високи концентрации, докато други компоненти на ДНК нямат това свойство.
През 1962 г., използвайки метода на рентгенова дифракция, беше възможно да се установи клетъчната структура на този гел. Оказа се, че се състои от четири остатъка от гуанин, които се свързват помежду си в кръг и образуват характерен квадрат. В центъра връзката се поддържа от метален йон (Na, K, Mg). Същите структури могат да се образуват в ДНК, ако съдържа много гуанин. Тези плоски квадратчета (G-квартети) са подредени, за да образуват сравнително стабилни, плътни структури (G-квадруплекси).
Четири отделни нишки на ДНК могат да бъдат преплетени в четириверижни комплекси, но това е по-скоро изключение. По-често единична верига нуклеинова киселина е просто свързана на възел, образувайки характерни удебелявания (например в краищата на хромозомите), или двуверижната ДНК образува локален четворка в някой регион, богат на гуанин.
Най -проучено е съществуването на квадруплекси в краищата на хромозомите - на теломери и в онкопромотори. Досега обаче не е известно пълно разбиране за локализацията на такава ДНК в човешки хромозоми.
Всички тези необичайни ДНК структури в линейна форма са нестабилни в сравнение с ДНК от В-форма. Въпреки това, ДНК често съществува в кръгова форма на топологичен стрес, когато има това, което се нарича суперсвиване. При тези условия лесно се образуват неканонични ДНК структури: Z-форми, кръстове и фиби, Н-форми, гуанинови квадруплекси и i-мотив.
- Свръхсвита форма - отбелязва се, когато се освобождава от клетъчното ядро, без да се повреди гръбнака на пентозофосфата. Той има формата на супер-усукани затворени пръстени. В свръхсвито състояние двойната спирала на ДНК е "усукана върху себе си" поне веднъж, тоест съдържа поне една суперспирала (приема формата на осмица).
- Отпуснатото състояние на ДНК се наблюдава с еднократно прекъсване (счупване на една верига). В този случай суперспиралите изчезват и ДНК приема формата на затворен пръстен.
- Линейна форма на ДНК - наблюдава се при счупване на две нишки на двойна спирала.
Третична структура на ДНК
Третична структура на ДНКсе образува в резултат на допълнително усукване в пространството на двуверижна молекула - нейното суперсвиване. Суперсвиването на молекулата на ДНК в еукариотни клетки, за разлика от прокариотите, се извършва под формата на комплекси с протеини.
Почти цялата еукариотна ДНК се намира в хромозомите на ядрата, само малко от нея се съдържа в митохондриите, в растенията и в пластидите. Основното вещество на хромозомите на еукариотни клетки (включително човешки хромозоми) е хроматин, състоящ се от двуверижна ДНК, хистонови и нехистонови протеини.
Хроматинови хистонови протеини
Хистоните са прости протеини, които съставляват 50% от хроматина. Във всички изследвани клетки на животни и растения бяха открити пет основни класа хистони: H1, H2A, H2B, H3, H4, различни по размер, аминокиселинен състави размера на таксата (винаги положителен).
Хистонът на бозайници Н1 се състои от единична полипептидна верига от около 215 аминокиселини; размерите на други хистони варират от 100 до 135 аминокиселини. Всички те са спирализирани и усукани в глобула с диаметър около 2,5 nm, съдържат необичайно голямо количество положително заредени аминокиселини лизин и аргинин. Хистоните могат да бъдат ацетилирани, метилирани, фосфорилирани, поли (ADP) -рибозилирани, а хистоните Н2А и Н2В са ковалентно свързани с убиквитин. Каква е ролята на такива модификации при формирането на структурата и изпълнението на функциите от хистони все още не е напълно изяснена. Предполага се, че това е тяхната способност да взаимодействат с ДНК и да осигурят един от механизмите за регулиране на действието на гените.
Хистоните взаимодействат с ДНК главно чрез йонни връзки (солни мостове), образувани между отрицателно заредени фосфатни групи на ДНК и положително заредени лизинови и аргининови остатъци от хистони.
Нехистонови хроматинови протеини
За разлика от хистоните, нехистоновите протеини са много разнообразни. Изолирани са до 590 различни фракции на ДНК-свързващи нехистонови протеини. Те се наричат още кисели протеини, тъй като киселинните аминокиселини преобладават в тяхната структура (те са полианиони). Специфичното регулиране на хроматиновата активност е свързано с различни нехистонови протеини. Например, ензимите, необходими за репликация и експресия на ДНК, могат временно да се свържат с хроматин. Други протеини, например, участващи в различни регулаторни процеси, се свързват с ДНК само в специфични тъкани или на определени етапи на диференциация. Всеки протеин е допълващ специфична ДНК нуклеотидна последователност (ДНК сайт). Тази група включва:
- семейство специфични за сайта протеини от цинков пръст. Всеки цинков пръст разпознава определено място, състоящо се от 5 двойки нуклеотиди.
- семейство специфични за сайта протеини - хомодимери. Фрагмент от такъв протеин в контакт с ДНК има структура спирала-завой-спирала.
- гел протеини с висока подвижност (HMG протеини) са група структурни и регулаторни протеини, които са постоянно свързани с хроматин. Те имат молекулно тегло по -малко от 30 kDa и се характеризират с високо съдържание на заредени аминокиселини. Поради ниското си молекулно тегло, HMG протеините са силно подвижни по време на електрофорезата с полиакриламиден гел.
- ензими на репликация, транскрипция и възстановяване.
С участието на структурни, регулаторни протеини и ензими, участващи в синтеза на ДНК и РНК, нуклеозомната нишка се превръща в силно кондензиран комплекс от протеини и ДНК. Получената структура е 10 000 пъти по -къса от оригиналната молекула на ДНК.

Хроматин
Хроматинът е комплекс от протеини с ядрена ДНК и неорганични вещества. По -голямата част от хроматина е неактивен. Той съдържа плътно опакована, кондензирана ДНК. Това е хетерохроматин. Разграничете конститутивния, генетично неактивен хроматин (сателитна ДНК), състоящ се от неизразени участъци, и факултативен - неактивен в редица поколения, но при определени обстоятелства, способни да се експресират.
Активният хроматин (еухроматин) е некондензиран, т.е. опаковани по -малко плътно. В различни клетки съдържанието му варира от 2 до 11%. В клетките на мозъка той е най -вече - 10-11%, в клетките на черния дроб - 3-4 и бъбреците - 2-3%. Отбелязва се активна транскрипция на еухроматин. Освен това неговата структурна организация позволява използването на същата генетична информация на ДНК, присъща на този видорганизъм, различно в специализирани клетки.
В електронен микроскоп изображението на хроматин прилича на мънисто: сферични удебелявания с размер около 10 nm, разделени от нишковидни мостове. Тези кълбовидни удебеления се наричат нуклеозоми. Нуклеозомата е структурна единица на хроматина. Всяка нуклеозома съдържа свръхсвързан сегмент от ДНК с дължина 146 базови двойки, рана с образуване на 1,75 леви завоя върху ядрото на нуклеозомата. Нуклеозомното ядро е хистонов октамер, състоящ се от хистони H2A, H2B, H3 и H4, по две молекули от всеки тип (фиг. 9), който прилича на диск с диаметър 11 nm и дебелина 5,7 nm. Петият хистон, H1, не е част от нуклеозомното ядро и не участва в процеса на навиване на ДНК върху хистоновия октамер. Той контактува с ДНК, където двойната спирала влиза и излиза от ядрото на нуклеозомата. Това са интеркортикални (линкерни) ДНК области, чиято дължина варира в зависимост от клетъчния тип от 40 до 50 нуклеотидни двойки. В резултат на това дължината на ДНК фрагмента, включен в нуклеозомата, също варира (от 186 до 196 двойки нуклеотиди).
Нуклеозомата съдържа около 90% от ДНК, останалата част е линкерът. Смята се, че нуклеозомите са фрагменти от "безшумен" хроматин и линкерът е активен. Нуклеозомите обаче могат да се разгънат и да станат линейни. Разгънатите нуклеозоми вече са активен хроматин. Така ясно се проявява зависимостта на функцията от структурата. Може да се предположи, че колкото повече хроматин е в състава на глобуларни нуклеозоми, толкова по -малко е активен. Очевидно е, че в различните клетки неравният дял на хроматина в покой е свързан с броя на такива нуклеозоми.
На електронно -микроскопични снимки, в зависимост от условията на изолация и степента на разтягане, хроматинът може да изглежда не само като дълга нишка с удебеления - „мъниста“ от нуклеозоми, но и като по -къса и по -плътна фибрила (влакно) с диаметър 30 nm, чието образуване се наблюдава по време на взаимодействие хистон Н1, свързан с линкерната област на ДНК и хистон Н3, което води до допълнително усукване на спирала от шест нуклеозоми на оборот с образуване на соленоид с диаметър 30 nm. В този случай хистонов протеин може да попречи на транскрипцията на редица гени и по този начин да регулира тяхната активност.
В резултат на гореописаните взаимодействия на ДНК с хистони, сегмент от двойната спирала на ДНК от 186 двойки основи със среден диаметър 2 nm и дължина 57 nm се трансформира в спирала с диаметър 10 nm и a дължина 5 nm. Последващото компресиране на тази спирала до влакно с диаметър 30 nm увеличава степента на кондензация с шест пъти.
В крайна сметка опаковането на дуплекс от ДНК с пет хистона води до 50-кратна кондензация на ДНК. Въпреки това, дори толкова висока степен на кондензация не може да обясни почти 50 000 - 100 000 пъти уплътняване на ДНК в метафазната хромозома. За съжаление, подробностите за по -нататъшното опаковане на хроматин до метафазната хромозома все още не са известни; следователно могат да се вземат предвид само общи характеристики на този процес.
Нива на уплътняване на ДНК в хромозоми
Всяка ДНК молекула е опакована в отделна хромозома. Човешките диплоидни клетки съдържат 46 хромозоми, които се намират в клетъчното ядро. Общата дължина на ДНК на всички хромозоми на клетка е 1,74 m, но диаметърът на ядрото, в което са опаковани хромозомите, е милиони пъти по -малък. Такова компактно опаковане на ДНК в хромозоми и хромозоми в клетъчното ядро се осигурява от различни хистонови и нехистонови протеини, които взаимодействат в специфична последователност с ДНК (виж по-горе). Уплътняването на ДНК в хромозомите дава възможност да се намалят нейните линейни размери с около 10 000 пъти - условно от 5 см до 5 микрона. Има няколко нива на уплътняване (фиг. 10).


- Двойната спирала на ДНК е отрицателно заредена молекула с диаметър 2 nm и дължина няколко cm.
- нуклеозомно ниво- хроматинът изглежда в електронен микроскоп като верига от "мъниста" - нуклеозоми - "на низ". Нуклеозомата е универсална структурна единица, която се намира както в еухроматина, така и в хетерохроматина, в междуфазното ядро и метафазните хромозоми.
Нуклеозомното ниво на уплътняване се осигурява от специални протеини - хистони. Осем положително заредени хистонови домена образуват сърцевината (ядрото) на нуклеозомата, около която е навита отрицателно заредената ДНК молекула. Това води до 7-кратно скъсяване, докато диаметърът се увеличава от 2 на 11 nm.
- ниво на соленоида
Соленоидното ниво на организация на хромозомите се характеризира с усукване на нуклеозомната нишка и образуването на по -дебели фибрили с диаметър 20-35 nm от нея - соленоиди или супербиди. Стъпката на соленоида е 11 nm; има около 6-10 нуклеозоми на ход. Опаковката на соленоиди се счита за по-вероятна от свръхбилната, според която хроматинов фибрил с диаметър 20-35 nm е верига от гранули или супербиди, всяка от които се състои от осем нуклеозоми. На нивото на соленоида линейният размер на ДНК се намалява с 6-10 пъти, диаметърът се увеличава до 30 nm.
- ниво на цикъл
Нивото на контура се осигурява от нехистонови сайт-специфични ДНК-свързващи протеини, които разпознават и се свързват със специфични ДНК последователности, образувайки бримки с приблизително 30-300 kb. Цикълът осигурява генна експресия, т.е. E. контурът е не само структурна, но и функционална формация. Съкращаването на това ниво се случва 20-30 пъти. Диаметърът се увеличава до 300 nm. На цитологични препарати могат да се видят примкови структури от типа "лампа-четка" в яйцеклетки земноводни. Тези бримки очевидно са суперсвити и представляват ДНК домейни, които вероятно отговарят на единиците за транскрипция и репликация на хроматин. Специфични протеини фиксират основите на бримките и евентуално някои от техните вътрешни области. Организацията на домейна във формата на бримка насърчава сгъването на хроматина в метафазните хромозоми в спираловидни структури от по-висок порядък.
- ниво на домейн
Нивото на домейна на хромозомната организация е недостатъчно проучено. На това ниво се отбелязва образуването на верижни домени - структури от нишки (фибрили) с дебелина 25-30 nm, които съдържат 60% протеин, 35% ДНК и 5% РНК, са практически невидими във всички фази на клетката цикъл с изключение на митозата и са донякъде произволно разпределени в клетъчното ядро. На цитологични препарати могат да се видят примкови структури от типа "лампа-четка" в яйцеклетки земноводни.
Домейните на бримката с тяхната основа са прикрепени към вътреядрената протеинова матрица в т. Нар. Вградени места за свързване, често наричани MAR / SAR последователности (MAR, от английския регион, свързан с матрицата; SAR, от английските региони за прикрепване на скелета) - ДНК фрагменти с няколкостотин базови двойки по дължина, които се характеризират с високо съдържание (> 65%) A / T базови двойки. Изглежда, че всеки домейн има един произход на репликация и функционира като самостоятелна суперсвъртена единица. Всеки цикличен домейн съдържа много транскрипционни единици, чието функциониране вероятно е координирано - целият домейн е или в активно, или в неактивно състояние.
На ниво домейн, в резултат на последователното опаковане на хроматин, линейните размери на ДНК намаляват с около 200 пъти (700 nm).
- хромозомно ниво
На хромозомно ниво кондензацията на профазната хромозома в метафазната хромозома се осъществява с уплътняване на домейни на контура около аксиалната рамка на нехистонови протеини. Това супер навиване е придружено от фосфорилиране на всички молекули H1 в клетката. В резултат на това метафазната хромозома може да бъде изобразена като плътно опаковани соленоидни бримки, навити в плътна спирала. Типичната човешка хромозома може да съдържа до 2600 бримки. Дебелината на такава структура достига 1400 nm (две хроматиди), докато молекулата на ДНК се скъсява 104 пъти, т.е. с 5 cm разтеглена ДНК до 5 μm.
Хромозомни функции
При взаимодействие с екстрахромозомни механизми, хромозомите осигуряват
- съхранение на наследствена информация
- използване на тази информация за създаване и поддържане на клетъчна организация
- регулиране на четенето на наследствена информация
- самоудвояване на генетичния материал
- прехвърляне на генетичен материал от майчината клетка към дъщерята.
Има доказателства, че при активиране на хроматинов регион, т.е. по време на транскрипцията първо от него се отстранява обратимо хистон Н1, а след това и хистонов октет. Това причинява декондензация на хроматин, последователен преход на 30-нанометров хроматинов фибрил в 10-нанометрова нишка и по-нататъшното му разгъване в области на свободна ДНК, т.е. загуба на нуклеозомна структура.
Екология на познанието. Наука и открития: Какво е животът? Този въпрос се превърна в движещата сила за развитието на генетиката (от гръцки genetikos - „свързан с раждането, произхода“) - науката за произхода на живота, която се фокусира върху молекулата на ДНК повече от 50 години.
Какво е живот? Този въпрос се превърна в движещата сила за развитието на генетиката (от гръцки genetikos - „свързан с раждането, произхода“) - науката за произхода на живота, която се фокусира върху молекулата на ДНК повече от 50 години.
Откритието, което обърна света с главата надолу
"Току -що открихме тайната на живота!" - така на 28 февруари 1953 г. Франсис Крик и Джеймс Уотсън обявяват откриването си на структурата на ДНК. Какво е донесло ново в науките за живота? Преди това беше известно, че ДНК е голяма молекула, в която информация за структурата и свойствата на живите същества се записва с помощта на "четирибуквената азбука". Но остава неясно как тази информация се предава от поколение на поколение и се материализира в тези структури и свойства, както и каква е пространствената структура на ДНК.
Решението на структурата на ДНК е помогнало на учените да разберат механизмите на нейното копиране и материализиране. ДНК се състои от две вериги, които се допълват (допълват) една към друга. Копирането на ДНК се дължи на добавянето на копие към всяка начална верига на ДНК, като върху шаблон, към него. Така от една двойна спирала на ДНК се получават две абсолютно еднакви двойни спирали, което е необходимо за запазване на генетичната информация по време на разделянето на майчината клетка на две дъщерни клетки. Принципът на матрицата също е в основата на постепенното материализиране на генетичната информация: върху една от нишките на ДНК се образува допълваща верига от друга информационна молекула - РНК, която от своя страна служи като матрица за синтеза на протеини, количеството и качеството от които зависи от структурата и свойствата на определен организъм.
Колко важно е това откритие за разбирането на мистерията на живота? От една страна, познаването на структурата на ДНК очевидно не е достатъчно, за да отговори на въпроса "какво е животът?" Но от друга страна, именно това откритие направи много древно и много важен въпросвърху връзката между потенциала и проявеното - чрез примера за връзка на информация за структурите и свойствата на един организъм със самите структури и свойства. И не само повдигна този въпрос, но и даде ключ към отговора на него. Този ключ е принципът на матрицата, принципът на взаимно допълване.
Пътят от ген до черта
Какво означава класическата фраза от учебника: „ДНК е носител на генетична информация“? Как генетичната информация е свързана със структурата на ДНК? Как информацията е въплътена в специфични свойства на организма? Ако вземем структурата на ДНК като отправна точка за генетична информация и след това следваме структурния модел, тогава пътят от гена до чертата ще изглежда така: всички свойства на организма са криптирани в последователността на ДНК; линейната структура на определен ген определя еднозначно линейната структура на съответния протеин, което от своя страна недвусмислено определя ролята на този протеин при формирането на определена черта.
С други думи, „ДНК поражда РНК; РНК ражда протеин, а протеинът ражда теб и мен ”(Ф. Крик). Ако това е вярно, тогава за да се промени тази или онази черта (например, за да се излекува заболяване, което има генетични корени), е достатъчно да се установи съответната последователност на ДНК секцията и да се коригира.
Но толкова ли е просто? Има ли достатъчно знания (въпреки че е безспорно правилно и необходимо!) За структурните съответствия по пътя от ген до черта, за да се разбере и възпроизведе този път?
Последните постижения в генетиката показват, че те са недостатъчни. През 2003 г., в рамките на проекта за човешки геном, линейната структура на човешката ДНК (и много други прости и сложни организми) беше напълно определена. Както каза един от учените, „всички букви, които съставят дебела книга, са прочетени, сега все още би било възможно да се разберат думите и тяхното значение“.
Оказа се, че има около 30 000 собствени гена (ДНК участъци, кодиращи протеини) при хората и това е само 1–3% от цялата ДНК! Същият брой гени в растението Arabidopsis taliana и рибата бухалка. Освен това 99% от човешките гени съвпадат с гените на мишката, тоест човекът има само 300 гена, които мишката няма. (Трудно е да си представим, че ние и мишките имаме същите 99% от признаците!)
Освен това. Оказа се, че недвусмислена връзка между ген и протеин съществува само в бактериите. А при хората е възможно да се образуват много протеини на базата на един ген (максималният известен брой различни протеини, кодирани от един ген днес, е 40 000!) И появата на много функции в един протеин. Оказва се, че пътят от потенциала към проявеното, от генетичната информация към чертата в никакъв случай не е линеен; че всяка черта е резултат от сложни взаимодействия между много гени и техните протеинови продукти; че самото понятие "ген" поради своята неяснота едва ли може да служи като "отправна точка" на този път.
Взаимодействие на структурите
Човешкото тяло се състои от 10 клетки до 14 -та степен. Всички те имат абсолютно еднаква ДНК, но се различават значително по форма, размер и своите задачи. Решението на този парадокс се крие в селективното четене на генетична информация. Във всяка клетка са активни само онези гени, от които тя се нуждае в момента.
Селективността се осигурява от специални регулаторни гени, които позволяват или забраняват четенето на информация от определен участък от ДНК. Генната активност също зависи от средата в пространството на клетъчното ядро. Промяна в околната среда, причинена от движението на самия ген или на някой от неговите съседи, може да промени неговата активност („изключване“ или „включване“ на гена).
Например, човешкият геном съдържа много потенциално опасни вирусни гени и протоонкогени (способни да причинят ракова дегенерация на клетка). Те могат да се държат доста спокойно за дълго време (и през целия си живот) и дори да работят за доброто на клетката, докато движението на тях самите или някой от околната среда не разкрие агресивни потенциали в тези гени. За щастие могат да възникнат и други движения, които да успокоят „бунтовника“ или да активират защитните механизми.
И така, носителят на генетична информация се е преместил от нивото на ген (специфичен участък от ДНК) към епигенетичното (от латинското „над“, „над“) ниво на взаимодействие на гените помежду си и с други структури на клетъчното ядро (99% от негенна ДНК и протеини). Да предположим, че науката дешифрира механизма на това взаимодействие. Ще доведе ли това до разкриването на тайната на живота? Животът само структура ли е? И ако не, струва ли си в търсене на улика за мистерията на живота да се ограничи до взаимодействието на структурите?
Кой гледа пазача?
Как от една клетка, в резултат на 46 деления, не се получава безформена маса от 1014 клетки, а много характерно тяло на всеки от нас? Постепенно удвоявайки се, клетките не само стават различни, но и образуват различни части от тялото правилното времеи на правилното място. Какво управлява организацията на клетките във времето и пространството?
Едно цяло, което е качествено по-голямо от обикновената сума от съставните му части-клетки. И това не противоречи на факта, че организмът се формира от една клетка - въпросът е какво за тази клетка олицетворява „волята на цялото“. Търсенето на такъв подреждащ фактор доведе до концепцията за морфогенетично поле в началото на 20 век. Негов основател е руският учен А.Г. Гурвич. Когато Гурвич работи върху теорията на полето, молекулата на ДНК се счита за неразделна част от хромозомите и не й се придава особено значение.
През 1944 г. ученият публикува своята работа "Теория на биологичното поле". Тази година се превърна в решаваща за цялата генетика, определяйки пътя на нейното развитие за няколко десетилетия напред. Фокусът на учените беше върху молекулата на ДНК, тъй като беше доказано, че именно тя играе водещата роля в предаването на наследствена информация. 1953 г. не беше далеч ... В резултат на това науката насочи цялото си внимание към структурата на ДНК, която всъщност започна да се отъждествява с информацията, която носи, а теорията за биологичното поле не беше на високо уважение. Но изследванията в тази област продължиха и през всичките тези години двата пътя за изучаване на тайните на живота вървяха паралелно ...
Преходът към новото хилядолетие промени баланса на силите в науките за живота. Все повече учени стигат до извода, че структурният ключ в познанието на живите е необходим, но не и достатъчен; че различните подходи не се изключват взаимно, а образуват единен път на научно изследване; че по същество структурният подход и теорията на полето се допълват.
Нека припомним: именно предположението за взаимното допълване на ДНК вериги стана ключът към дешифрирането на нейната структура, а откриването на самата 1953 г. стана възможно благодарение на взаимното допълване на усилията на представители на различни области на науката - физици, химици, биолози . Може би единната наука от новото хилядолетие не само най -накрая ще съгласува различните научни подходи (например структурни и теренни), но и ще се обърне към плодовете на „ненаучния“ начин на изучаване на тайните на живота - хилядолетната мъдрост на човечеството - "ненаучно", тъй като този път се корени в онези времена, когато изобщо нямаше наука.
Обръщането към източниците на древна мъдрост може да даде на науката ключовете към вратата, зад която се крие тайната. Но за да се случи това, така различни начинизнанието трябва да се „пресече“ някъде. Едно от тези „кръстовища“ може да бъде концепцията за формообразуващо поле (биологично, морфогенетично, информационно), което е нараснало въз основа на съвременната наука и се връща към явленията, разглеждани в древните източници.
Последните казват, че човек се състои от няколко тела или принципи, които не са отделни, независими части, но се проникват и взаимно се оформят; че видимото, плътно, физическо тяло е проводник, носител на по -фини тела, които с негова помощ се проявяват във физическия свят и взаимодействат с него; че "сборката" - образуването на физическо тяло от елементи на физическата материя - се контролира от "най -плътното" от тези тела, астралното тяло -прототип (план, матрица).
Съвременната наука знае, че всяка новообразувана клетка участва във формирането на организъм според индивидуална „инструкция“ (активни или включени гени) и че родителят и потомците на тази клетка може да имат напълно различни „инструкции“. Но какво и как ще координира превключването на отделни „програми“ за развитие на милиарди клетки, все още не е ясно.
Теорията на биологичното поле приема, че координацията е функция на цялото, което е самото поле, матрица или модел; че всяка новообразувана клетка с помощта на собствен генетичен апарат е свързана с един -единствен „план“ на развитието на организма, получава оттам индивидуални „инструкции“ и ги изпълнява в рамките на собствената си програма за поведение.
Оказва се, че генетичният апарат на клетката се състои от поне три функционални блока: приемаща „антена“, „контролен панел“ за генна активност и „изпълнителна“ част - гени, отговорни за образуването на специфични протеини. Припомнете си, че гените представляват само 1-2% от цялата клетъчна ДНК. В останалите 98-99% от ДНК вече са открити структури, свързани с втория „контролен блок“. И какво играе ролята на "антена"? Къде се извършва „срещата на два свята“ - информационното поле и генетичните структури, които въплъщават тази информация в конкретно физическо тяло?
Защо не приемем, че ДНК също играе ролята на антена, способна да улавя, трансформира и предава сигнали с вълново естество? Спиралната структура на "молекулата на живота" (много технически антени имат формата на спирала) и нейните свойства като способността да провеждат електрически ток, възможността за резонансно възбуждане на надлъжни трептения под действието на радиовълни, както и като способност за генериране на лазерна светлина след предварително "изпомпване" ".
Ако ДНК може да работи за получаване на информация, която осигурява жизнената активност на клетките, тогава не е необходимо постоянно да съхранява тази информация в своята структура. Както например човешкият мозък, за да контролира успешно системите за поддържане на живота на тялото, не е задължително да бъде „вместилище“ на ума, но е достатъчно да играе ролята на посредник между съзнанието и тяло: възприема информацията от равнината на съзнанието и я „превежда“ на езика на телесния контрол.
И е ясно защо физическото тяло страда в случай на увреждане на ДНК структурата (или мозъчните структури). В края на краищата всеки знае, че в случай на неизправност в поне една част от телевизора, изображението на екрана му е силно изкривено и ако телевизорът е лишен от антената или е изключен от мрежата, нищо няма да се появи на екран изобщо.
ДНК е връзката между "модела" на физическото тяло и неговото конкретно въплъщение. Мозъкът е посредник между ума и тялото. Разумът свързва живота и формата на неговото проявление и позволява на живота, затворен във форма, да опознае себе си. С помощта на този прекрасен инструмент човек има възможност да изследва света около себе си и да намери в него ключовете за познаване на своя вътрешен свят. Така се ражда единен път, водещ до познаване на тайната на живота. Защото човекът е най -голямата мистерия - мистерията на връзката между земята и небето.публикувани
P.S. И помнете, просто като промените консумацията си - заедно променяме света! © еконет
Присъединете се към нас на
След откриването на принципа на молекулярна организация на вещество като ДНК през 1953 г., молекулярната биология започва да се развива. Освен това, в процеса на изследване, учените установиха как ДНК се рекомбинира, нейния състав и как работи нашият човешки геном.
Най -сложните процеси протичат на молекулярно ниво всеки ден. Как е структурирана молекулата на ДНК, от какво се състои? И каква роля играят молекулите на ДНК в клетката? Нека поговорим подробно за всички процеси, протичащи в двойната верига.
Какво е наследствена информация?
И така, откъде започна всичко? Още през 1868 г. те са открити в ядрата на бактерии. И през 1928 г. Н. Колцов излага теорията, че именно в ДНК цялата генетична информация за жив организъм е криптирана. Тогава Дж. Уотсън и Ф. Крик откриват модел на сега известната ДНК спирала през 1953 г., за което заслужено са получили признание и награда - Нобелова награда.
Какво все пак е ДНК? Това вещество се състои от 2 обединени нишки, по -точно спирали. Секция от такава верига с определена информация се нарича ген.

ДНК съхранява цялата информация за това кои протеини ще се образуват и в какъв ред. ДНК макромолекулата е материален носител на невероятно обемна информация, която се записва чрез строга последователност от отделни градивни елементи - нуклеотиди. Общо има 4 нуклеотида, те се допълват взаимно химически и геометрично. Този принцип на взаимно допълване или допълване в науката ще бъде описан по -късно. Това правило играе ключова роля при кодирането и декодирането на генетична информация.
Тъй като веригата на ДНК е невероятно дълга, няма повторения в тази последователност. Всяко живо същество има своя уникална нишка ДНК.
ДНК функции
Функциите включват съхранение на наследствена информация и предаването й на потомството. Без тази функция геномът на вида не би могъл да бъде запазен и развит в продължение на хилядолетия. Организмите, които са претърпели големи генни мутации, е по -вероятно да не оцелеят или да загубят способността си да произвеждат потомство. Така възниква естествената защита от дегенерацията на вида.

Друга съществена функция е внедряването на съхранена информация. Клетката не може да направи нито един жизненоважен протеин без инструкциите, съхранявани в двойна верига.
Състав на нуклеинова киселина
Сега вече е надеждно известно от какво се състоят самите нуклеотиди - градивните елементи на ДНК. Те се състоят от 3 вещества:
- Ортофосфорна киселина.
- Азотна основа. Пиримидинови основи - които имат само един пръстен. Те включват тимин и цитозин. Пуринови основи, които съдържат 2 пръстена. Това са гуанин и аденин.
- Захароза. Като част от ДНК - дезоксирибоза, В РНК - рибоза.
Броят на нуклеотидите винаги е равен на броя на азотните основи. В специални лаборатории се разцепва нуклеотид и от него се изолира азотна основа. Така се изучават индивидуалните свойства на тези нуклеотиди и възможните мутации в тях.
Нива на организация на наследствената информация
Има 3 нива на организация: генно, хромозомно и геномно. Цялата информация, необходима за синтеза на нов протеин, се съдържа в малка част от веригата - гена. Тоест генът се счита за най -ниското и най -простото ниво на кодиране на информация.

Гените от своя страна се сглобяват в хромозоми. Благодарение на тази организация на носителя на наследствен материал групи герои се редуват според определени закони и се предават от едно поколение на друго. Трябва да се отбележи, че в тялото има невероятно много гени, но информацията не се губи дори когато се рекомбинира многократно.
Има няколко вида гени:
- според функционалното им предназначение има 2 вида: структурни и регулаторни последователности;
- според влиянието върху процесите, протичащи в клетката, те се разграничават: супервитални, смъртоносни, условно смъртоносни гени, както и мутаторни и антимутаторни гени.
Гените са разположени по протежение на хромозомата в линеен ред. В хромозомите информацията не е случайно фокусирана, има определен ред. Има дори карта, показваща позициите или локусите на гените. Например, известно е, че хромозома номер 18 кодира данни за цвета на очите на детето.
Какво е геном? Това е името на целия набор от нуклеотидни последователности в клетката на тялото. Геномът характеризира целия вид, а не отделен индивид.
Какъв е генетичният код на човека?
Факт е, че целият огромен потенциал човешко развитиезаложени още в периода на зачеването. Цялата наследствена информация, необходима за развитието на зиготата и растежа на детето след раждането, е кодирана в гените. Части от ДНК са най -основните носители на наследствена информация.
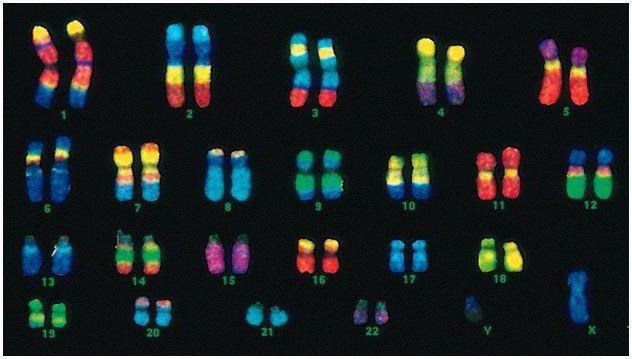
Човек има 46 хромозоми или 22 соматични двойки плюс по една хромозома за определяне на пола от всеки родител. Този диплоиден набор от хромозоми кодира цялостния външен вид на човек, неговите умствени и физически способности и предразположение към заболяване. Соматични хромозомивъншно неразличими, но носят различна информация, тъй като единият от тях е от бащата, а другият от майката.
Мъжкият код се различава от женския в последната двойка хромозоми - XY. Женският диплоиден комплект е последната двойка, XX. Мъжете получават една Х хромозома от биологичната си майка и след това тя се предава на дъщерите им. Половата хромозома Y се предава на синове.
Човешките хромозоми се различават значително по размер. Например, най -малката двойка хромозоми е # 17. А най -голямата двойка е 1 и 3.
Диаметърът на двойна спирала при хората е само 2 nm. ДНК е усукана толкова плътно, че се вписва в малкото ядро на клетката, въпреки че тя ще бъде дълга до 2 метра, ако е навита. Дължината на спиралата е стотици милиони нуклеотиди.
Как се предава генетичният код?
И така, каква роля играят молекулите на ДНК в клетката по време на деленето? Гените - носители на наследствена информация - се намират във всяка клетка на тялото. За да предадат своя код на дъщерния организъм, много същества разделят своята ДНК на 2 еднакви спирали. Това се нарича репликация. В процеса на репликация ДНК се размотва и специални "машини" допълват всяка верига. След като генетичната спирала се раздвои, ядрото и всички органели започват да се делят, а след това и цялата клетка.
Но човек има различен процес на трансфер на гени - сексуален. Чертите на бащата и майката са смесени, новият генетичен код съдържа информация и от двамата родители.
Съхранението и предаването на наследствена информация е възможно поради сложната организация на ДНК спиралата. В края на краищата, както казахме, структурата на протеините е кодирана в гени. Веднъж създаден по време на зачеването, този код ще се копира през целия си живот. Кариотипът (личен набор от хромозоми) не се променя по време на обновяването на клетката на органа. Предаването на информация се осъществява с помощта на сексуални гамети - мъжки и женски.
Само вирусите, съдържащи една нишка РНК, не са в състояние да предадат своята информация на потомството си. Следователно, за да се размножават, те се нуждаят от човешки или животински клетки.
Реализиране на наследствена информация
В ядрото на клетките постоянно се появяват важни процеси... Цялата информация, записана в хромозомите, се използва за изграждане на протеини от аминокиселини. Но ДНК веригата никога не напуска ядрото, така че тук е необходима помощта на друго важно съединение = РНК. Именно РНК е в състояние да проникне в ядрената мембрана и да взаимодейства с веригата ДНК.
Чрез взаимодействието на ДНК и 3 типа РНК се реализира цялата кодирана информация. На какво ниво е прилагането на наследствена информация? Всички взаимодействия се осъществяват на нуклеотидно ниво. Messenger РНК копира част от ДНК веригата и пренася това копие в рибозомата. Тук започва синтезът на нова молекула от нуклеотиди.
За да може иРНК да копира необходимата част от нишката, спиралата се разгъва и след приключване на процеса на прекодиране се възстановява отново. Освен това този процес може да се осъществи едновременно от 2 страни на 1 хромозома.
Принципът на взаимно допълване
Те се състоят от 4 нуклеотида - аденин (А), гуанин (G), цитозин (С), тимин (Т). Те са свързани чрез водородни връзки съгласно правилото за допълване. Работите на Е. Чаргаф помогнаха за установяването на това правило, тъй като ученият забеляза някои модели в поведението на тези вещества. Е. Чаргаф открива, че моларното съотношение на аденина към тимина е равно на единица. И по същия начин съотношението гуанин към цитозин винаги е равно на едно.
Въз основа на неговата работа генетиката формира правило за взаимодействието на нуклеотидите. Правилото за допълване гласи, че аденинът се комбинира само с тимин, а гуанин с цитозин. По време на декодирането на спиралата и синтеза на нов протеин в рибозомата, такова правило за редуване помага за бързото намиране незаменима аминокиселинакойто е прикрепен към транспортната РНК.
РНК и нейните видове
Какво е наследствена информация? нуклеотиди в двойна верига на ДНК. Какво е РНК? Каква е нейната работа? РНК или рибонуклеинова киселина помага да се извлече информация от ДНК, да се декодира и, въз основа на принципа на комплементарност, да се създадат протеини, необходими за клетките.
Общо са изолирани 3 вида РНК. Всеки от тях стриктно изпълнява своята функция.
- Информационен (мРНК), или иначе се нарича матрица. Той отива направо в центъра на клетката, в ядрото. Открива в една от хромозомите необходимия генетичен материал за изграждане на протеин и копира една от страните на двойната верига. Копирането се случва отново според принципа на взаимно допълване.
- ТранспортЕ малка молекула, която има нуклеотидни декодери от едната страна и аминокиселини, съответстващи на основния код от другата страна. Задачата на тРНК е да я достави до „работилницата“, тоест до рибозомата, където синтезира необходимата аминокиселина.
- рРНК - рибозомна.Той контролира количеството произведен протеин. Състои се от 2 части - аминокиселина и пептиден сайт.
Единствената разлика в декодирането е, че РНК няма тимин. Вместо тимин има урацил. Но след това, в процеса на протеинов синтез, с тРНК, той все още правилно задава всички аминокиселини. Ако има някакви грешки при декодирането на информация, тогава възниква мутация.
Възстановяване на повредена ДНК молекула
Процесът на поправяне на повредена двойна нишка се нарича ремонт. По време на процеса на възстановяване повредените гени се отстраняват.

След това необходимата последователност от елементи се възпроизвежда точно и се отрязва на същото място на веригата, откъдето е извлечена. Всичко това се случва благодарение на специалното химикали- ензими.
Защо се появяват мутации?
Защо някои гени започват да мутират и спират да изпълняват своята функция - съхраняване на жизненоважна наследствена информация? Това се дължи на грешка при декодиране. Например, ако аденинът случайно се замени с тимин.
Има и хромозомни и геномни мутации. Хромозомните мутации възникват, когато части от наследствена информация се загубят, дублират или дори се прехвърлят и интегрират в друга хромозома.

Геномните мутации са най -сериозните. Тяхната причина е промяна в броя на хромозомите. Тоест, когато вместо чифт - диплоиден набор, в кариотипа присъства триплоиден набор.
Най -известният пример за триплоидна мутация е синдромът на Даун, при който личен набор от хромозоми е 47. При такива деца се образуват 3 хромозоми на мястото на 21 -та двойка.
Известна е и мутация като полиплоидия. Но полиплоидията се среща само в растенията.
ДНК молекулата се състои от две нишки, които образуват двойна спирала. Структурата му е дешифрирана за първи път от Франсис Крик и Джеймс Уотсън през 1953 г.
Първоначално молекулата на ДНК, състояща се от двойка нуклеотидни вериги, усукани една около друга, повдигна въпроса защо има точно тази форма. Учените наричат този феномен допълващ се, което означава, че само определени нуклеотиди могат да бъдат разположени в неговите нишки един срещу друг. Например, аденинът винаги е срещу тимина, а гуанинът е срещу цитозин. Тези нуклеотиди на молекулата на ДНК се наричат комплементарни.
Това е схематично изобразено, както следва:
Т - А
C - G
Тези двойки образуват химическа нуклеотидна връзка, която определя реда, в който са подредени аминокиселините. В първия случай е малко по -слаб. Връзката между C и G е по -силна. Некомплементарните нуклеотиди не образуват двойки помежду си.
Относно структурата
И така, структурата на молекулата на ДНК е специална. Той има такава форма по някаква причина: факт е, че броят на нуклеотидите е много голям и е необходимо много място за настаняване на дълги вериги. Поради тази причина спиралното усукване е присъщо на веригите. Това явление се нарича спирализация, което позволява на нишките да се скъсят около пет до шест пъти.
Тялото използва някои молекули от този вид много активно, други рядко. Последните, в допълнение към спирализацията, също се подлагат на такава "компактна опаковка" като супер навиване. И тогава дължината на молекулата на ДНК намалява 25-30 пъти.

Какво представлява "опаковането" на молекула?
В процеса на суперсвиване се включват хистонови протеини. Те имат структурата и вида на резбова макара или пръчка. Върху тях се навиват спираловидни нишки, които веднага стават „компактно опаковани“ и заемат малко място. Когато се наложи да се използва тази или онази нишка, тя се размотава от намотка, например, на хистонов протеин, и спиралата се размотава в две паралелни вериги. Когато молекулата на ДНК е в това състояние, от нея могат да се отчетат необходимите генетични данни. Има обаче едно условие. Получаването на информация е възможно само ако структурата на молекулата на ДНК не е изкривена. Наличните за четене хромозоми се наричат еухроматини и ако са суперсипирализирани, това вече са хетерохроматини.
Нуклеинова киселина
Нуклеиновите киселини, подобно на протеините, са биополимери. Основната функция е съхранението, внедряването и предаването на наследствена (генетична информация). Те са два вида: ДНК и РНК (дезоксирибонуклеинова и рибонуклеинова). Мономери в тях са нуклеотиди, всеки от които съдържа остатък от фосфорна киселина, пет въглеродна захар (дезоксирибоза / рибоза) и азотна основа. ДНК кодът включва 4 вида нуклеотиди - аденин (А) / гуанин (G) / цитозин (С) / тимин (Т). Те се различават по съдържащата се в тях азотна основа.
В молекулата на ДНК броят на нуклеотидите може да бъде огромен - от няколко хиляди до десетки и стотици милиони. Такива гигантски молекули могат да се видят през електронен микроскоп. В този случай ще бъде възможно да се види двойна верига от полинуклеотидни нишки, които са свързани помежду си чрез водородни връзки на азотните основи на нуклеотидите.

Изследвания
В хода на изследванията учените са установили, че видовете ДНК молекули в различните живи организми се различават. Установено е също, че гуанинът от една верига може да се свърже само с цитозин, а тиминът - с аденин. Разположението на нуклеотидите на една нишка строго съответства на паралелната. Поради тази допълняемост на полинуклеотидите, молекулата на ДНК е способна на дублиране и самовъзпроизвеждане. Но първо, комплементарните вериги се разминават под въздействието на специални ензими, които разрушават сдвоени нуклеотиди, а след това синтезът на липсващата верига започва във всеки от тях. Това се дължи на Голям бройсвободни нуклеотиди във всяка клетка. В резултат на това вместо "родителската молекула" се образуват две "дъщерни" молекули, идентични по състав и структура и ДНК кодът става оригиналният. Този процес е предшественик на клетъчното делене. Той осигурява прехвърлянето на всички наследствени данни от майчините клетки към дъщерните клетки, както и към всички следващи поколения.
Как се чете генният код?
Днес се изчислява не само масата на молекулата на ДНК - можете да откриете и по -сложни данни, които преди това не бяха достъпни за учените. Например, можете да прочетете информация за това как тялото използва собствената си клетка. Разбира се, първоначално тази информация е кодирана и има формата на някаква матрица и затова тя трябва да бъде транспортирана до специален носител, който е РНК. Рибонуклеиновата киселина е в състояние да проникне в клетката през ядрената мембрана и да прочете кодираната информация вътре. По този начин РНК е носител на скрити данни от ядрото до клетката и се различава от ДНК по това, че съдържа рибоза вместо дезоксирибоза и урацил вместо тимин. В допълнение, РНК е едноверижна.

Синтез на РНК
Дълбокият анализ на ДНК показа, че след като РНК напусне ядрото, тя навлиза в цитоплазмата, където може да бъде включена като шаблон в рибозоми (специални ензимни системи). Въз основа на получената информация те могат да синтезират подходящата последователност от протеинови аминокиселини. Рибозомата научава от триплетния код какъв вид органично съединение трябва да бъде прикрепено към образуващата се протеинова верига. Всяка аминокиселина има свой специфичен триплет, който я кодира.
След като образуването на веригата приключи, тя придобива специфична пространствена форма и се превръща в протеин, способен да изпълнява своите хормонални, изграждащи, ензимни и други функции. За всеки организъм той е генен продукт. Именно от него се определят всякакви качества, свойства и прояви на гени.

Гени
На първо място, процесите на секвениране са разработени, за да се получи информация за това колко гени има структурата на молекулата на ДНК. И въпреки че изследванията позволяват на учените да постигнат голям напредък по този въпрос, все още не е възможно да се знае точния им брой.
Преди няколко години се предполагаше, че молекулите на ДНК съдържат приблизително 100 хиляди гени. Малко по -късно цифрата намалява до 80 хиляди, а през 1998 г. генетиците обявяват, че само 50 хиляди гени присъстват в една ДНК, което е само 3% от цялата дължина на ДНК. Но последните заключения на генетиците бяха изумени. Сега те твърдят, че геномът включва 25-40 хиляди от тези единици. Оказва се, че само 1,5% от хромозомната ДНК е отговорна за кодиращите протеини.
Изследванията не спряха дотук. Паралелен екип от специалисти по генно инженерство установи, че броят на гените в една молекула е точно 32 хиляди. Както можете да видите, все още е невъзможно да се получи окончателен отговор. Има твърде много противоречия. Всички изследователи разчитат само на собствените си резултати.
Имаше ли еволюция?
Въпреки факта, че няма доказателства за еволюцията на молекулата (тъй като структурата на молекулата на ДНК е крехка и с малки размери), учените направиха едно предложение. Въз основа на лабораторни данни те изразиха версия на следното съдържание: в началния етап на появата си молекулата изглеждаше като прост самовъзпроизвеждащ се пептид, който включваше до 32 аминокиселини, открити в древните океани.
След самовъзпроизвеждане, благодарение на силите на естествения подбор, молекулите придобиха способността да се предпазват от влиянието на външни елементи. Те започнаха да живеят по -дълго и да се възпроизвеждат в голям брой. Молекулите, които се озоваха в липидния пикочен мехур, получиха всички шансове за самовъзпроизвеждане. В резултат на поредица от последователни цикли липидните мехурчета придобиват формата на клетъчни мембрани и едва след това - всички известни частици... Трябва да се отбележи, че днес всяка част от молекулата на ДНК е сложна и ясно функционираща структура, всички характеристики на която все още не са напълно проучени от учените.

Съвременен свят
Наскоро израелски учени са разработили компютър, който може да извършва трилиони операции в секунда. Днес това е най -бързата кола на Земята. Цялата тайна е, че иновативното устройство се захранва от ДНК. Професорите казват, че в краткосрочен план такива компютри дори ще могат да генерират енергия.
Специалисти от института Weizmann в Реховот (Израел) преди година обявиха създаването на програмируем молекулен компютър, състоящ се от молекули и ензими. Те замениха силициевите микрочипове с тях. Към момента екипът продължава да се движи напред. Сега само една молекула ДНК може да предостави на компютъра необходимите данни и да осигури необходимото гориво.
Биохимичните „нанокомпютри“ не са измислица, те вече съществуват в природата и се проявяват във всяко живо същество. Но често те не се контролират от хора. Човек все още не може да оперира с генома на което и да е растение, за да изчисли, да речем, числото "pi".
Идеята за използване на ДНК за съхранение / обработка на данни за пръв път удари светлите умове на учените през 1994 г. Тогава молекулата беше използвана за решаване на проста математическа задача. Оттогава редица изследователски групи предлагат различни проекти, свързани с ДНК компютри. Но тук всички опити се основават само на енергийната молекула. Не можете да видите такъв компютър с просто око; той изглежда като прозрачен разтвор на вода в епруветка. В него няма механични части, а само трилиони биомолекулни устройства - и това е само в една капка течност!
Човешка ДНК
За каква човешка ДНК хората са узнали през 1953 г., когато учените за първи път са успели да демонстрират пред света двуверижен модел на ДНК. За това Кърк и Уотсън получиха Нобелова награда, тъй като това откритие стана фундаментално през 20 -ти век.
С течение на времето, разбира се, те доказаха, че структурирана човешка молекула може да изглежда не само като в предложената версия. След по-подробен анализ на ДНК бяха открити А-, В- и лявата форма на Z-. Форма А- често е изключение, тъй като се образува само при липса на влага. Но това е възможно само при лабораторни изследвания, за естествената среда е ненормално, в жива клетка такъв процес не може да настъпи.
Формата B е класическа и е известна като двойна дясна верига, но Z-формата не само е усукана в обратната посока, наляво, но има и по-зигзагообразен вид. Учените са идентифицирали и G-квадруплексната форма. В неговата структура няма 2, а 4 нишки. Според генетиците тази форма се среща в онези райони, където има излишно количество гуанин.

Изкуствена ДНК
Изкуствена ДНК вече съществува днес, която е идентично копие на истинската; той перфектно повтаря структурата на естествената двойна спирала. Но за разлика от първичния полинуклеотид, в изкуствения има само два допълнителни нуклеотида.
Тъй като дублирането е създадено въз основа на информация, получена в хода на различни изследвания на реална ДНК, то може да се копира, самовъзпроизвежда и еволюира. Експертите работят около създаването на такава изкуствена молекула от около 20 години. Резултатът е невероятно изобретение, което може да използва генетичния код по същия начин като естествената ДНК.
Към четирите налични азотни основи генетиката добави още две, които са създали по метода на химическата модификация на естествените основи. За разлика от естествената ДНК, изкуствената ДНК е доста къса. Той съдържа само 81 двойки основи. Той обаче също се възпроизвежда и развива.
Получена репликация на молекула изкуствено, се осъществява благодарение на полимеразната верижна реакция, но засега това не се случва независимо, а чрез намесата на учените. Те независимо добавят необходимите ензими към споменатата ДНК, като я поставят в специално приготвена течна среда.
Краен резултат
Процесът и крайният резултат от развитието на ДНК могат да бъдат повлияни от различни фактори, например мутации. Това налага изследване на проби от материя, така че резултатът от анализите да е надежден и надежден. Пример за това е тестът за бащинство. Но е добра новина, че такива инциденти като мутация са редки. Независимо от това пробите от материя винаги се проверяват отново, за да се получи по -точна информация въз основа на анализа.

Растителна ДНК
Благодарение на високотехнологичното секвениране (HTS) е направена революция в областта на геномиката - възможно е и извличането на ДНК от растенията. Разбира се, получаването на висококачествено молекулно тегло на ДНК от растителен материал причинява известни трудности поради големия брой копия на ДНК митохондрии и хлоропласти, както и високо нивополизахариди и фенолни съединения. За да изолираме структурата, която разглеждаме, в този случай се използват различни методи.
Водородна връзка в ДНК
Водородната връзка в молекулата на ДНК е отговорна за електромагнитното привличане, създадено между положително заредения водороден атом, който е прикрепен към електроотрицателен атом. Това диполно взаимодействие не отговаря на критерия химическа връзка... Но може да се реализира междумолекулно или в различни части на молекулата, тоест вътремолекулно.

Водородният атом е прикрепен към електроотрицателния атом, който е донор на тази връзка. Електроотрицателният атом може да бъде азот, флуор, кислород. Той - чрез децентрализация - привлича електронен облак от водородното ядро и прави водородния атом зареден (частично) положително. Тъй като размерът на Н е малък в сравнение с други молекули и атоми, зарядът също е малък.
ДНК декодиране
Преди да декодират молекула на ДНК, учените първо вземат огромен брой клетки. За най -точната и успешна работа те се нуждаят от около милион. Резултатите, получени в хода на изследването, постоянно се сравняват и записват. Днес декодирането на генома вече не е рядкост, а достъпна процедура.

Разбира се, дешифрирането на генома на една клетка е неподходящо упражнение. Данните, получени в хода на такива изследвания, не представляват интерес за учените. Но е важно да се разбере, че всички съществуващи понастоящем методи за декодиране, въпреки тяхната сложност, не са достатъчно ефективни. Те ще позволят само 40-70% от ДНК да бъде прочетена.
Преподавателите от Харвард обаче наскоро обявиха начин за дешифриране на 90% от генома. Техниката се основава на добавянето на праймерни молекули към изолираните клетки, с помощта на които започва репликацията на ДНК. Но дори и този метод не може да се счита за успешен; той все още трябва да бъде подобрен, преди да бъде открито използван в науката.
ДНК (дезоксирибонуклеинова киселина) се отнася (заедно с РНК), които са полимери, или по -скоро полинуклеотиди (мономер - нуклеотид).
ДНК е отговорна за съхраняването и предаването на генетичния код по време на клетъчното делене. Чрез молекулите на ДНК се реализират наследствеността и променливостта. Всички видове РНК се синтезират върху ДНК. Освен това, различни видове РНК съвместно осигуряват синтеза на клетъчни протеини, тоест те реализират генетична информация.
В еукариотните клетки огромното количество ДНК се намира в ядрото, където те образуват комплекси със специални протеини, което води до образуването на хромозоми. В прокариотни клетки има една голяма кръгла (или линейна) ДНК молекула (също в комплекс с протеини). В допълнение, еукариотните клетки имат собствена ДНК в митохондриите и хлоропластите.
В случая на ДНК всеки нуклеотид се състои от 1) азотна основа, която може да бъде аденин, гуанин, цитозин или тимин, 2) дезоксирибоза, 3) фосфорна киселина.
Последователността на нуклеотидите в ДНК веригата определя първичната структура на молекулата. ДНК се характеризира с вторична структура на молекулата под формата на двойна спирала (най-често с дясна ръка). В този случай две ДНК вериги са свързани помежду си чрез водородни връзки, образувани между комплементарни азотни бази.
Аденинът допълва тимина, а гуанинът допълва цитозина. Между аденин и тимин се образуват две водородни връзки, а три между гуанин и цитозин. По този начин гуанинът и цитозинът са свързани помежду си малко по -силно (въпреки че водородните връзки по принцип са слаби). Броят на връзките се определя от структурните особености на молекулите.
Аденинът и гуанинът са пурини и се състоят от два пръстена. Тиминът и цитозинът се отнасят до пиримидинови основи, състоящи се от един пръстен. По този начин, между гръбнаците (състоящи се от редуващи се дезоксирибоза и фосфорна киселина) на две нишки на ДНК, за всяка двойка нуклеотиди от различни нишки, винаги има три пръстена (тъй като пуринът с два пръстена винаги е допълващ само за определен единичен пръстен пиримидин). Това ви позволява да поддържате ширината между нишките на молекулата на ДНК еднаква по цялата дължина (около 2,3 nm).
В един завой на спиралата има около 10 нуклеотида. Дължината на един нуклеотид е приблизително 0,34 nm. Дължината на молекулите на ДНК обикновено е огромна, надхвърляща милиони нуклеотиди. Следователно, за да се побере по -компактно в клетъчното ядро, ДНК претърпява различни степени на „суперсвиване“.
При четене на информация от ДНК (тоест синтезиране на РНК върху нея, този процес се нарича транскрипция), настъпва деспирализация (процес на обратна спирализация), двете вериги се разминават под действието на специален ензим. Водородните връзки са слаби, поради което отделянето и последващото омрежване на веригите се случва с ниска консумация на енергия. РНК се синтезира върху ДНК съгласно същия принцип на комплементарност. Само вместо тимин в РНК, урацилът е комплементарен на аденина.
Генетичен код, написана върху молекули на ДНК, се състои от триплети (последователности от три нуклеотида), които представляват една аминокиселина (протеин мономер). Повечето от ДНК обаче не кодират протеин. Значението на такива части от молекулата е различно, в много отношения не е напълно разбрано.
Преди клетъчното делене винаги има удвояване на количеството ДНК. Този процес се нарича репликация... Той е полуконсервативен по природа: веригите на една молекула ДНК се разминават и всяка има своя собствена нова допълваща се верига. В резултат на това от една двуверижна ДНК молекула се получават две двуверижни ДНК, идентични с първата.
В ДНК полинуклеотидните вериги са многопосочни, тоест където една верига има 5 "край (остатъкът от фосфорна киселина е прикрепен към петия въглероден атом на дезоксирибоза), а другата има 3" край (без въглерод от фосфорна киселина) .
По време на репликацията и транскрипцията синтезът винаги протича в посока от 5 "края към 3", тъй като новите нуклеотиди могат да се прикрепят само към свободния 3 "въглероден атом.
Структурата и ролята на ДНК като вещество, отговорно за наследствената информация, е изяснено през 40-50-те години на ХХ век. През 1953 г. Д. Уотсън и Ф. Крик идентифицират двуверижната структура на ДНК. По -рано Е. Чаргаф установява, че в ДНК количеството на тимина винаги съответства на аденина, а количеството на гуанин - на цитозина.
Подобни статии
-
Романи за тийнейджъри (тийнейджърски любовни книги)
Никога не съм мислил за утре, докато не се събудих от предозиране в болницата. Не исках да се събудя. Но те ме спасиха. - Имали сте трансплантация на сърце. Защо го направиха? Сега сърцето на някой друг бие в гърдите ми и аз ...
-
Най -мъдрите цитати на Омар Хаям за живота и любовта
Който е присадил роза на нежна любов към порязванията на сърцето - не е живял напразно! И този, който слушаше Бога със сърцето си, И този, който пиеше хмела на земната наслада! О, горко, горко на сърцето, където няма изгаряща страст. Където няма любов към мъките, където няма мечти за щастие. Ден без ...
-
Най -красивите реплики от песни
Всички умираме, но не всички живеем.Жените искат любов, стабилност, честност. По принцип като всички хора. Животът е игра, основното е да не преигравате. Хап и млъкни. Забравете за мен, забравете, аз съм вашето табу. Нищо не може да бъде върнато. Съжалявам, ти мен ...
-
Вярно ли е, че инженерите правят оборудване, което се разрушава нарочно с течение на времето?
Трябва да започнем с факта, че всяко оборудване рано или късно ще се повреди - това определено е факт. Рядко се случва повреда на оборудването след определен експлоатационен живот, но такова оборудване съществува и обикновено е скъпо. Несъмнено производителите се интересуват от ...
-
Джим Рейнър - история на героите
Космическата опера StarCraft 2 продължава. Във втората част на трилогията, зерг расата излиза на преден план. Главният герой в Сърцето на рояка е Сара Кериган, един от ключовите герои във Вселената. Не всеки познава добре тази дама ...
-
Съвременният младежки речник: основни тенденции
Речникът на всеки език се актуализира и обогатява постепенно. Заемането на чужди думи играе съществена роля в това. Все по -често английските думи се използват в руската реч по отношение на: науката (астронавт, наблюдение, ...