Білок: будова і функції. Властивості білків. Органічні речовини. Амінокислоти. Білки, що повторюються ланки білків називаються
Органічні речовини.До складу живих організмів, крім неорганічних, входять також різноманітні органічні речовини. Органічні речовини живих істот утворені, перш за все, чотирма хімічними елементами, званими біогенними: Вуглецем, воднем, киснем і азотом. У складі білків до цих елементів додається сірка, а в нуклеїнових кислотах - фосфор.
Різноманіття органічних речовин в значній мірі визначається вуглецем. Цей елемент завдяки унікальним властивостям становить хімічну основу життя. Він може утворювати ковалентні зв'язки з багатьма атомами і їх групами, утворюючи ланцюжки, каблучки, складові скелет різних за складом, будовою, довжині і формі органічних молекул. З них в свою чергу, утворюються складні хімічні сполуки, Що розрізняються за будовою і функціями. Основна причина різноманітності органічних молекул - це не стільки відміну складових їх атомів, скільки різний порядок їх розміщення в молекулі.
Поняття про біополімерах.В живому організмі органічні речовини представлені або невеликими, з відносно низькою молекулярною масою молекулами, або великими макромолекулами. До низькомолекулярних сполук відносяться амінокислоти, цукри, органічні кислоти, спирти, вітаміни та ін.
Білки, полісахариди і нуклеїнові кислоти в більшості своїй є структурами з великою молекулярною масою. Тому їх називають макромолекулами (Від грец. макрос - великий). Так, молекулярна маса більшості білків становить від 5000 до 1 000000. Високомолекулярні органічні сполуки - білки, нуклеїнові кислоти, полісахариди, молекули яких складаються з великої кількості однакових або різних за хімічною будовою повторюваних ланок, називаються биополимерами(Від грец. биос - життя та поліс - численний). прості молекули, Із залишків яких складаються біополімери, називаються мономерами. Мономерами білків є амінокислоти, полісахаридів - моносахариди, нуклеїнових кислот - нуклеотиди. Макромолекули складають близько 90% сухої маси клітини.
У цьому розділі розглянуто всі три класи макромолекул і їх мономерні ланки. До розгляду додані ліпіди - молекули, як правило, значно дрібніші, ніж біополімери, але також виконують функції в організмі.
Особливу групу органічних речовин складають біологічно активні речовини: ферменти, гормони, вітаміни та ін. Вони різноманітні за будовою; впливають на обмін речовин і перетворення енергії.
У клітинах різних груп організмів зміст певних органічних сполук різний. Наприклад, в клітинах тварин переважають білки і жири, а в клітинах рослин - вуглеводи. Однак в різних клітинах певні органічні сполуки виконують схожі функції.
Білки.В живих організмах серед макромолекул за своїм функціональним значенням провідна роль належить білкам. Білки в багатьох організмах переважають і кількісно. Так, в організмі тварин вони становлять 40-50%, в організмі рослин - 20 - 35% їх сухої маси. Білки - це гетерополімери, мономерами яких є амінокислоти.
Амінокислоти - «цеглинки» білкових молекул. амінокислоти - органічні сполуки, що містять одночасно аміногрупу (-NН), для якої характерні основні властивості, і карбоксильну групу (-СООН) з кислотними властивостями. Аміногрупа і карбоксильная групи пов'язані з одним і тим же атомом вуглецю (рис.). За цією ознакою всі амінокислоти подібні між собою. У більшій частині белокобразующіх амінокислот є одна карбоксильна група і одна аміногрупа; ці амінокислоти називаються нейтральними.
Частина молекули, званої радикалом (R) у різних амінокислот має різну будову (рис.). Радикал у різних амінокислот може бути неполярних або полярним (зарядженим або незарядженим), гідрофобним або гідрофільним, що і надає білків певні властивості. Крім нейтральних, існують основні амінокислоти- з більш ніж однієї аминогруппой, а також кислі амінокислоти- з більш ніж однієї карбоксильної групою. Наявність додаткової амино- або гідроксильної групи впливає на властивості радикала. Всі властивості радикалів амінокислот відіграють визначальну роль у формуванні просторової структури білка.
Загальне число відомих амінокислот близько 200, а в освіті природних білків бере участь тільки 20 відовт. Такі амінокислоти називаються белокобразующімі (Таблиця 2; в таблиці наведені повне та скорочене назви амінокислот, не для запам'ятовування).
Таблиця 2. Основні амінокислоти і їх скорочене позначення
Рослини і бактерії можуть синтезувати всі необхідні їм амінокислоти з первинних продуктів фотосинтезу. Людина і тварини не здатні синтезувати всі амінокислоти, тому так звані незамінні амінокислоти вони повинні отримувати в готовому вигляді разом з їжею.
Незамінними амінокислотами для людини є: лізин, валін, лейцин, ізолейцин, треонін, фенілаланін, триптофані метіонін; для дітей незамінними є також аргініні гистидин. Білки їжі, що містять всі незамінні амінокислоти, називаються повноцінними, на відміну від неповноцінних, В яких відсутні ті чи інші незамінні амінокислоти.
Наявність в одній амінокислоті і основний, і кислотної груп обумовлює їх амфотерность і високу реактивність. аміногрупа
(-NH 2) однієї амінокислоти здатна взаємодіяти з карбоксильною групою (-СООН) іншої амінокислоти з виділенням молекули води. Утворюється при цьому молекула являє собою дипептид (Рис.), А зв'язок СО-NН- називається пептидного. На одному кінці молекули дипептиду знаходиться вільна аміногрупа, а на іншому карбоксильная група. Завдяки цьому дипептид може приєднувати до себе інші амінокислоти, утворюючи олігопептиди. Якщо таким чином з'єднується багато амінокислот (більше десяти), то утворюється довгий ланцюг - поліпептид.
Пептиди грають важливу роль в організмі. Багато олиго- і поліпептиди є гормонами, антибіотиками, токсинами.
До олігопептиди відносяться, наприклад, гормони гіпофіза окситоцин і вазопресин, а також брадикінін (пептид болю) і деякі опіати ( «природні наркотики» людини), що виконують функцію знеболювання. Регулярне вживання Вживання наркотиків дуже небезпечно, воно руйнує опіатних систему організму, тому наркоман без дози наркотиків відчуває сильний біль - «ломку». До олігопептиди відносяться деякі антибіотики, наприклад, граміцидин S.
Гормони (інсулін, адренокортикотропний гормон та ін.), Антибіотики (граміцидин А), токсини (дифтерійний токсин) також є поліпептидами.
Поліпептидні ланцюги бувають дуже довгими і включають найрізноманітніші комбінації амінокислот. Поліпептиди, в молекулу яких входить від 50 до кількох тисяч амінокислотних залишків з молекулярною масою понад 6000, називаються білками.
Кожен конкретний білок характеризується строго постійним складом і послідовністю амінокислотних залишків.
Рівні організації білкової молекули.Молекули білків можуть приймати різні просторові форми – конформації, які представляють собою чотири рівні їх організації (рис.)
Ланцюжок з безлічі амінокислотних залишків, сполучених пептидними зв'язками є первинну структуру білкової молекули. Це найбільш важлива структура, так як вона визначає його форму, властивості і функції. На основі первинної структури створюються інші види структур. Саме ця структура закодована в молекулі ДНК. Кожен індивідуальний білок організму має унікальну первинну структуру. Всі молекули конкретного індивідуального білка (наприклад, альбуміну) мають однакове чергування амінокислотних залишків, що відрізняє альбумін від будь-якого іншого індивідуального білка. Різноманіття первинної структури визначається складом, кількістю і порядком проходження амінокислотних залишків у поліпептидному ланцюзі.
вторинна структура білків виникає в результаті утворення водневих зв'язків між атомом водню NH-груп і атомом кисню CO-груп різних амінокислотних залишків поліпептидного ланцюга. Поліпептидний ланцюг при цьому закручується в спіраль. Хоча водневі зв'язку слабкі, але завдяки значній кількості вони забезпечують стабільність цієї структури. Повністю спіральну конфігурацію мають молекули білка кератину. Це структурний білок волосся, вовни, кігтів, пір'я і рогів; він входить до складу зовнішнього шару шкіри хребетних. Крім кератину спіральна вторинна структура характерна для фібрилярних (ниткоподібних) білків, таких як міозин, фібриноген, колаген.
Вторинна структура білка, крім спіралі, може бути представлена \u200b\u200bскладчастим шаром. У складчатом шарі кілька поліпептидних ланцюгів (або ділянок одного поліпептидного ланцюга) лежать паралельно, утворюючи плоску конфігурацію, складену на зразок гармошки (рис. Б6). Вторинну структуру в формі складчастого шару має, наприклад, білок натурального, що становить основну масу шовкового волокна, що виділяється шелкоотделітельнимі залозами гусениці шовкопряда при сплетінні коконів.
третинна структура створюється S-S зв'язками ( «дисульфідними містками») між залишками цистеїну (амінокислота, що містить сірку), а також водневими, іонними та іншими взаємодіями. Третинної структурою визначаються специфічність білкових молекул, їх біологічна активність. Третинну структуру мають такі білки, як міоглобін (білок, що знаходиться в м'язах, бере участь у створенні запасів кисню), трипсин (фермент, який розщеплює білки в кишечнику).
У деяких випадках кілька поліпептидних ланцюгів з третинної структурою об'єднуються в єдиний комплекс, при цьому утворюється четвертичная структура. У ній білкові субодиниці не пов'язані ковалентно, а міцність забезпечується взаємодією слабких міжмолекулярних сил. Наприклад, четвертичная структура характерна для білка гемоглобіну, що складається з чотирьох білкових субодиниць і небілкової частини - гема.
s 1. Що таке білки? 2. Яке будова білків? 3. Що таке амінокислоти? 4. Яким чином амінокислоти з'єднуються в поліпептидний ланцюг? 5. Які рівні структурної організації білків існують? 6. які хімічні зв'язки обумовлюють різні рівні структурної організації білкових молекул? 7. Є три види амінокислот А.В.С. Скільки варіантів поліпептидних ланцюгів, що складаються з п'яти амінокислот, можна побудувати? Чи будуть поліпептиди мати однакові властивості?
Короткий опис:
Фрагмент підручника: Біологічна хімія з вправами і завданнями: підручник / за ред. чл.-кор. РАМН С.Є. Северина. М .: ГЕОТАР-Медіа, 2011. - 624 с .: іл. МОДУЛЬ 1: БУДОВА, ВЛАСТИВОСТІ І ФУНКЦІЇ БІЛКІВ
МОДУЛЬ 1: БУДОВА, ВЛАСТИВОСТІ І ФУНКЦІЇ БІЛКІВ
структура модуля | теми |
Модульна одиниця 1 | 1.1. Структурна організація білків. Етапи формування нативної конформації білків 1.2. Основи функціонування білків. Ліки як ліганди, що впливають на функцію білків 1.3. Денатурація білків і можливість їх спонтанної ренатіваціі |
Модульна одиниця 2 | 1.4. Особливості будови і функціонування олігомерних білків на прикладі гемоглобіну 1.5. Підтримка нативной конформації білків в умовах клітини 1.6. Різноманіття білків. Сімейства білків на прикладі імуноглобулінів 1.7. Фізико-хімічні властивості білків і методи їх розподілу |
Модульна одиниця 1 СТРУКТУРНА ОРГАНІЗАЦІЯ мономірним БІЛКІВ І ОСНОВИ ЇХ ФУНКЦІОНУВАННЯ
Цілі вивчення Вміти:
1. Використовувати знання про особливості структури білків і залежності функцій білків від їх структури для розуміння механізмів розвитку спадкових і набутих протеінопатій.
2. Пояснювати механізми лікувальної дії деяких ліків як лігандів, що взаємодіють з білками і змінюють їх активність.
3. Використовувати знання про будову і конформаційної лабільності білків для розуміння їх структурно-функціональної нестійкості і схильності до денатурації в умовах, що змінюються.
4. Пояснювати застосування денатуруючих агентів в якості засобів для стерилізації медичного матеріалу та інструментів, а також в якості антисептиків.
знати:
1. Рівні структурної організації білків.
2. Значення первинної структури білків, що визначає їх структурний і функціональний різноманіття.
3. Механізм формування в білках активного центру і його специфічну взаємодію з лігандом, що лежить в основі функціонування білків.
4. Приклади впливу екзогенних лігандів (ліків, токсинів, отрут) на конформацію і функціональну активність білків.
5. Причини і наслідки денатурації білків, фактори, що викликають денатурацію.
6. Приклади використання денатуруючих факторів в медицині як антисептики і засобів для стерилізації медичних інструментів.
ТЕМА 1.1. СТРУКТУРНА ОРГАНІЗАЦІЯ БІЛКІВ. ЕТАПИ ФОРМУВАННЯ нативному
конформація БІЛКІВ
білки - це гетерополімерние молекули (Тобто складаються з різноманітних мономерів). Мономерами білків є 20 видів α-амінокислот, з'єднані між собою пептидними зв'язками.
Набір і порядок з'єднання амінокислот у білку визначається будовою відповідних генів в ДНК индивидумов. Кожен білок відповідно до його специфічною структурою виконує властиву йому функцію. Набір білків даного організму ( протеом) Визначає його фенотипічні особливості, а також наявність спадкових хвороб або схильність до їх розвитку.
1. Амінокислоти, що входять до складу білків. Пептидний зв'язок.
Білки - гетерополімери, побудовані з мономерів - 20 α-амінокислот.
Загальна формула амінокислот представлена \u200b\u200bнижче.

Амінокислоти розрізняються за будовою, розмірами, фізико-хімічними властивостями радикалів, приєднаних до α-вуглецевого атома. Функціональні групи амінокислот визначають особливості властивостей різних α-амінокислот. Зустрічаються в α-амінокислотах радикали можна розділити на кілька груп:
пролин,на відміну від інших 19 мономерів білків, що не амінокислота, а імінокіслота, радикал в проліном пов'язаний як з α-вуглецевим атомом, так і з іміногрупи

Амінокислоти розрізняються за розчинністю у воді.Це пов'язано зі здатністю радикалів взаємодіяти з водою (гідратованих).
До гідрофільнимвідносяться радикали, що містять аніонні, катіонні і полярні незаряджені функціональні групи.
До гідрофобнимвідносяться радикали, що містять метильние групи, аліфатичні ланцюги або цикли.
2. Пептидні зв'язки з'єднують амінокислоти в пептиди.При синтезі пептиду α-карбоксильная група однієї амінокислоти взаємодіє з α-аміногрупою іншої амінокислоти з утворенням пептидного зв'язку:
Білки являють собою поліпептиди, тобто лінійні полімери α-амінокислот, з'єднаних пептидним зв'язком (рис. 1.1.)
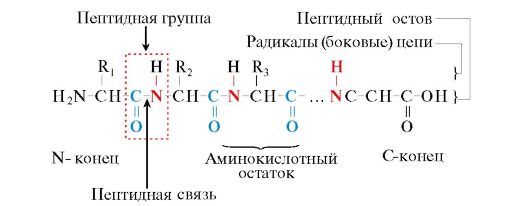 Мал. 1.1. Терміни, які використовуються при описі будови пептидів
Мал. 1.1. Терміни, які використовуються при описі будови пептидів
Мономери амінокислот, що входять до складу поліпептидів, називаються амінокислотними залишками.Ланцюг повторюваних груп - NH-CH-CO- утворює пептидний кістяк.Амінокислотний залишок, який має вільну α-аміногрупу, називається N-кінцевим, а має вільну α-карбоксильної групи - С-кінцевим. Пептиди записують і читають з N-кінця до С-кінця.
Пептидний зв'язок, утворена іміногрупи пролина, відрізняється від інших пептидних зв'язків: у атома азоту пептидної групи відсутній водень,
замість нього є зв'язок з радикалом, в результаті одна сторона циклу включається в пептидний кістяк:
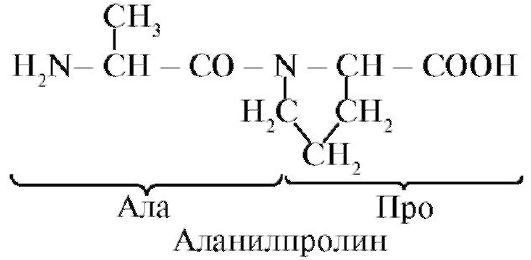
пептиди розрізняються амінокислотним складом, Кількістю амінокислот і порядком з'єднання амінокислот, наприклад, Сер-Ала-Гли-Гіс і Гіс-Гли-Ала-Сер - два різних пептиду.
Пептидні зв'язку дуже міцні, і для їх хімічного неферментативного гідролізу потрібні жорсткі умови: аналізований білок гідролізують в концентрованій соляній кислоті при температурі близько 110 ° С протягом 24 годин. У живій клітині пептидні зв'язку можуть розриватися за допомогою протеолітичних ферментів,званих протеазамиабо пептідгідролази.
3. Первинна структура білків.Амінокислотні залишки в пептидних ланцюгах різних білків чергуються не випадковим чином, а розташовані в певному порядку. Лінійна послідовність або порядок чергування амінокислотних залишків у поліпептидному ланцюзі називається первинною структурою білка.
Первинна структура кожного індивідуального білка закодована в молекулі ДНК (в ділянці, званому геном) і реалізується в ході транскрипції (переписування інформації на мРНК) і трансляції (синтез первинної структури білка). Отже, первинна структура білків індивідуального людини - спадково передається від батьків дітям інформація, яка визначає особливості будови білків даного організму, від яких залежить функція наявних білків (рис. 1.2.).
 Мал. 1.2. Взаємозв'язок між генотипом і конформацией білків, що синтезуються в організмі індивіда
Мал. 1.2. Взаємозв'язок між генотипом і конформацией білків, що синтезуються в організмі індивіда
Кожен з приблизно 100 000 індивідуальних білків в організмі людини має унікальнупервинну структуру. У молекулах одного типу білка (наприклад, альбуміну) однакове чергування амінокислотних залишків, що відрізняє альбумін від будь-якого іншого індивідуального білка.
Послідовність амінокислотних залишків в пептидного ланцюга можна розглядати як форму запису інформації. Ця інформація визначає просторову укладання лінійної пептидного ланцюга в більш компактну тривимірну структуру, яка називається конформациейбілка. Процес формування функціонально активної конформації білка носить назву фолдінг.
4. Конформація білків.Вільне обертання в пептидному кістяку можливо між атомом азоту пептидної групи і сусіднім α-вуглецевим атомом, а також між α-вуглецевим атомом і вуглецем карбонільної групи. Внаслідок взаємодії функціональних груп амінокислотних залишків первинна структура білків може набувати більш складні просторові структури. У глобулярних білках розрізняють два основних рівня укладання конформації пептидних ланцюгів: вториннуі третинну структури.
Вторинна структура білків- це просторова структура, що формується в результаті утворення водневих зв'язків між функціональними групами -С \u003d О і - NH- пептидного остова. При цьому пептидная ланцюг може набувати регулярні структури двох типів: α-спіраліі β-структури.
В α-спіраліводневі зв'язки утворюються між атомом кисню карбонільної групи і воднем амідного азоту 4-й від нього амінокислоти; бічні ланцюга амінокислотних залишків
розташовуються по периферії спіралі, не беручи участь в утворенні вторинної структури (рис. 1.3.).
Об'ємні радикали або радикали, що несуть однакові заряди, перешкоджають формуванню α-спіралі. Залишок пролина, що має кільцеву структуру, перериває α-спіраль, так як через відсутність водню у атома азоту в пептидного ланцюга неможливо утворити водневий зв'язок. Зв'язок між азотом і α-вуглецевим атомом входить до складу циклу пролина, тому пептидний кістяк в цьому місці набуває вигин.
β-Структураформується між лінійними областями пептидного остова однієї поліпептидного ланцюга, утворюючи при цьому складчасті структури. Поліпептидні ланцюги або їх частини можуть формувати паралельніабо антипаралельні β-структури.У першому випадку N- і С-кінці взаємодіючих пептидних ланцюгів збігаються, а в другому - мають протилежний зміст (рис. 1.4).
Мал. 1.3. Вторинна структура білка - α-спіраль
 Мал. 1.4. Паралельні і антипаралельні β-складчасті структури
Мал. 1.4. Паралельні і антипаралельні β-складчасті структури
β-структури позначені широкими стрілками: А - антипаралельними β-структура. Б - Паралельні β-складчасті структури
У деяких білках β-структури можуть формуватися за рахунок утворення водневих зв'язків між атомами пептидного остова різних поліпептидних ланцюгів.
У білках також зустрічаються області з нерегулярною вторинноїструктурою, до яких відносять вигини, петлі, повороти поліпептидного остова. Вони часто розташовуються в місцях, де змінюється напрямок пептидного ланцюга, наприклад, при формуванні паралельної β-складчастої структури.
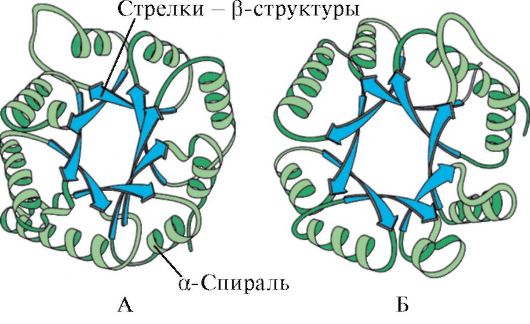
За наявністю α-спіралей і β-структур глобулярні білки можуть бути розділені на чотири категорії.
Мал. 1.5. Вторинна структура міоглобіну (А) і β-ланцюга гемоглобіну (Б), що містять вісім α-спіралей

 Мал. 1.6. Вторинна структура тріозофосфатізомерази і домена піруваткінази
Мал. 1.6. Вторинна структура тріозофосфатізомерази і домена піруваткінази
 Мал. 1.7. Вторинна структура константного домену імуноглобуліну (А) і ферменту супероксиддисмутази (Б)
Мал. 1.7. Вторинна структура константного домену імуноглобуліну (А) і ферменту супероксиддисмутази (Б)
В четверту категоріювключені білки, що мають в своєму складі незначну кількість регулярних вторинних структур. До таких білків можна віднести невеликі, багаті цистеїном білки або металопротеїни.
Третинна структура білка- тип конформації, що утворюється за рахунок взаємодій між радикалами амінокислот, які можуть перебувати на значній відстані один від одного в пептидного ланцюга. Більшість білків при цьому формують просторову структуру, що нагадує глобулу (глобулярні білки).
Так як гідрофобні радикали амінокислот мають тенденцію до об'єднання за допомогою так званих гідрофобних взаємодійі міжмолекулярних ван дер Ваальсових сил, всередині білкової глобули утворюється щільне гидрофобное ядро. Гідрофільні іонізовані і неіонізовані радикали в основному розташовуються на поверхні білка і визначають його розчинність в воді.
 Мал. 1.8. Типи зв'язків, що виникають між радикалами амінокислот при формуванні третинної структури білка
Мал. 1.8. Типи зв'язків, що виникають між радикалами амінокислот при формуванні третинної структури білка
1 - іонна зв'язок- виникає між позитивно і негативно зарядженими функціональними групами;
2 - воднева зв'язок- виникає між гидрофильной незарядженою і будь-який інший гидрофильной групою;
3 - гідрофобні взаємодії- виникають між гідрофобними радикалами;
4 - дисульфідний зв'язок- формується за рахунок окислення SH-груп залишків цистеїну і їх взаємодії один з одним
Гідрофільні амінокислотні залишки, які опинилися всередині гідрофобного ядра, можуть взаємодіяти один з одним за допомогою іоннихі водневих зв'язків(Рис. 1.8).
Іонні і водневі зв'язку, а також гідрофобні взаємодії відносяться до числа слабких: їх енергія не набагато перевищує енергію теплового руху молекул при кімнатній температурі. Конформація білка підтримується за рахунок виникнення безлічі таких слабких зв'язків. Так як атоми, з яких складається білок, знаходяться в постійному русі, то можливий розрив одних слабких зв'язків і утворення інших, що призводить до невеликих переміщень окремих ділянок поліпептидного ланцюга. Це властивість білків змінювати конформацію в результаті розриву одних і утворення інших слабких зв'язків називається конформационной лабільністю.
В організмі людини функціонують системи, що підтримують гомеостаз- постійність внутрішнього середовища в певних допустимих для здорового організму межах. В умовах гомеостазу невеликі зміни конформації не порушують загальну структуру і функцію білків. Функціонально активна конформація білка називається нативной конформацией.Зміна внутрішнього середовища (наприклад, концентрації глюкози, іонів Са, протонів і т.д.) призводить до зміни конформації і порушення функцій білків.
Третинна структура деяких білків стабілізована дисульфідними зв'язками,утворюються за рахунок взаємодії -SH груп двох залишків
 Мал. 1.9. Освіта дисульфідній зв'язку в молекулі білка
Мал. 1.9. Освіта дисульфідній зв'язку в молекулі білка
цистеїну (рис. 1.9). Більшість внутрішньоклітинних білків не має в третинної структурі ковалентних дисульфідних зв'язків. Їх наявність характерно для секретується клітиною білків, що забезпечує їх велику стабільність в позаклітинних умовах. Так, дисульфідні зв'язки є в молекулах інсуліну і імуноглобулінів.
інсулін- білковий гормон, що синтезується в β-клітинах підшлункової залози і секретується в кров у відповідь на підвищення концентрації глюкози в крові. У структурі інсуліну є дві дисульфідні зв'язки, що з'єднують поліпептидні А- і В-ланцюга, і одна дисульфідний зв'язок всередині А-ланцюга (рис. 1.10).
 Мал. 1.10. Дисульфідні зв'язки в структурі інсуліну
Мал. 1.10. Дисульфідні зв'язки в структурі інсуліну
5. Супервторічная структура білків.У різних по первинній структурі і функціям білках іноді виявляються подібні поєднання і взаємне розташування вторинних структур,які називаються супервторічной структурою. Вона займає проміжне положення між вторинної і третинної структурами, оскільки це специфічне поєднання елементів вторинної структури при формуванні третинної структури білка. Супервторічние структури мають специфічні назви, такі як «α-спіраль-поворот-а-спіраль», «лейцінових застібка блискавка», «цинкові пальці» і ін. Такі супервторічние структури характерні для ДНК-зв'язуючих білків.
«Лейцінових застібка-блискавка».Цей вид супервторічной структури використовується для з'єднання двох білків. На поверхні взаємодіючих білків є α-спіральні ділянки, що містять не менше чотирьох залишків лейцину. Лейцінових залишки в α-спіралі розташовуються через шість амінокислот один від іншого. Так як кожен виток α-спіралі містить 3,6 амінокислотних залишку, радикали лейцину знаходяться на поверхні кожного другого витка. Лейцінових залишки α-спіралі одного білка можуть взаємодіяти з лейцінових залишками іншого білка (гідрофобні взаємодії), поєднуючи їх разом (рис. 1.11.). Багато ДНК зв'язуючі білки функціонують в складі олігомерних комплексів, де окремі субодиниці зв'язуються один з одним «лейцінових застібками».
 Мал. 1.11. «Лейцінових застібка-блискавка» між α-спіральними ділянками двох білків
Мал. 1.11. «Лейцінових застібка-блискавка» між α-спіральними ділянками двох білків
Прикладом таких білків можуть служити гістони. гістони- ядерні білки, до складу яких входить велика кількість позитивно заряджених амінокислот - аргініну і лізину (до 80%). Молекули гістонів об'єднуються в олігомерні комплекси, що містять вісім мономерів за допомогою «лейцінових застібок», незважаючи на значний однойменний заряд цих молекул.
«Цинкову палець»- варіант супервторічной структури, характерний для ДНК-зв'язуючих білків, має вигляд витягнутого фрагмента на поверхні білка і містить близько 20 амінокислотних залишків (рис. 1.12). Форму «витягнутого пальця» підтримує атом цинку, пов'язаний з радикалами чотири амінокислот - двох залишків цистеїну і двох - гистидина. У деяких випадках замість залишків гістидину знаходяться залишки цистеїну. Два близько лежачі залишку цистеїну відокремлені від двох інших залишків Гісілі Ціспоследовательностью, що складається приблизно з 12 амінокислотних залишків. Ця ділянка білка утворює α-спіраль, радикали якої можуть специфічно зв'язуватися з регуляторними ділянками великий борозенки ДНК. Специфічність зв'язування індивідуального
 Мал. 1.12. Первинна структура ділянки ДНК-зв'язуючих білків, які формують структуру «цинкового пальця» (буквами позначені амінокислоти, що входять до складу цієї структури)
Мал. 1.12. Первинна структура ділянки ДНК-зв'язуючих білків, які формують структуру «цинкового пальця» (буквами позначені амінокислоти, що входять до складу цієї структури)
регуляторного ДНК-зв'язуючого білка залежить від послідовності амінокислотних залишків, розташованих в області «цинкового пальця». Такі структури містять, зокрема, рецептори стероїдних гормонів, Що беруть участь в регуляції транскрипції (зчитування інформації з ДНК на РНК).
ТЕМА 1.2. ОСНОВИ ФУНКЦІОНУВАННЯ БІЛКІВ. ЛІКИ ЯК ліганд, ЩО ВПЛИВАЮТЬ НА ФУНКЦІЮ БІЛКІВ
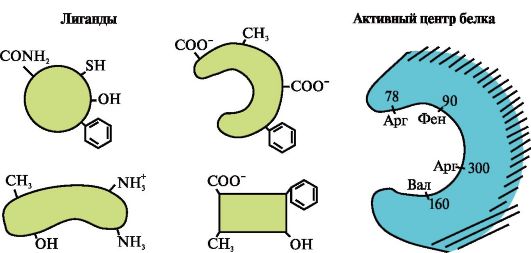
1. Активний центр білка і його взаємодію з лігандом.У процесі формування третинної структури на поверхні функціонально активного білка, зазвичай в поглибленні, утворюється ділянка, сформований радикалами амінокислот, далеко стоять один від одного в первинній структурі. Ця ділянка, що має унікальну будову для даного білка і здатний специфічно взаємодіяти з певною молекулою або групою схожих молекул, називається центром зв'язування білка з лігандом або активним центром. Лігандами називаються молекули, які взаємодіють з білками.
висока специфічністьвзаємодії білка з лігандом забезпечується комплементарностью структури активного центру структурі ліганду.
комплементарність- це просторове і хімічне відповідність взаємодіючих поверхонь. Активний центр повинен не тільки просторово відповідати входить в нього ліганду, а й між функціональними групами радикалів, що входять в активний центр, і лігандом повинні утворитися зв'язку (іонні, водневі, а також гідрофобні взаємодії), які утримують ліганд в активному центрі (рис. 1.13 ).
 Мал. 1.13. Комплементарна взаємодія білка з лігандом
Мал. 1.13. Комплементарна взаємодія білка з лігандом
Деякі ліганди, приєднуючись до активного центру білка, виконують допоміжну роль у функціонуванні білків. Такі ліганди називаються кофакторами, а білки, що мають в своєму складі небілкової частина, - складними білками(На відміну від простих білків, що складаються тільки з білкової частини). Небілкова частина, міцно поєднана з білком, носить назву простетичної групи.Наприклад, в складі міоглобіну, гемоглобіну і цитохромов міститься міцно прикріплена до активного центру простетичної група - гем, що містить іон заліза. Складні білки, що містять гем, називаються гемопротеинов.
При приєднанні до білків специфічних лігандів проявляється функція цих білків. Так, альбумін - найважливіший білок плазми крові - виявляє свою транспортну функцію, приєднуючи до активного центру гідрофобні ліганди, такі як жирні кислоти, білірубін, деякі ліки і ін. (Рис. 1.14)
Лігандами, взаємодіючими з тривимірною структурою пептидного ланцюга, можуть бути не тільки низькомолекулярні органічні та неорганічні молекули, але і макромолекули:
ДНК (розглянуті вище приклади з ДНК-зв'язуючими білками);
полісахариди;
 Мал. 1.14. Взаємозв'язок генотипу і фенотипу
Мал. 1.14. Взаємозв'язок генотипу і фенотипу
Унікальна первинна структура білків людини, закодована в молекулі ДНК, в клітинах реалізується у вигляді унікальної конформації, структури активного центру і функцій білків
У цих випадках білок дізнається певну ділянку лиганда, співрозмірний і комплементарний центру зв'язування. Так на поверхні гепатоцитів є білки-рецептори до гормону інсуліну, що має також білкове будова. Взаємодія інсуліну з рецептором викликає зміна його конформації і активації сигнальних систем, що призводять до запасання в гепатоцитах поживних речовин після їжі.
Таким чином, в основі функціонування білків лежить специфічна взаємодія активного центру білка з лігандом.
2. Доменна структура і її роль у функціонуванні білків.Довгі поліпептидні ланцюга глобулярних білків часто складаються в кілька компактних, відносно незалежних областей. Вони мають самостійну третинну структуру, що нагадує таку у глобулярних білків, і називаються доменами.Завдяки доменної структурі білків легше формується їх третинна структура.
У доменних білках центри зв'язування з лігандом часто розташовуються між доменами. Так, трипсин - протеолітичний фермент, який виробляється екзокринної частиною підшлункової залози і необхідний для перетравлення білків їжі. Він має двухдоменное будова, а центр зв'язування трипсину з його лігандом - харчовим білком - розташовується в борозенці між двома доменами. В активному центрі створюються умови, необхідні для ефективного зв'язування специфічного ділянки харчового білка і гідролізу його пептидних зв'язків.
Різні домени в білку при взаємодії активного центру з лігандом можуть переміщатися одна відносно одної (рис. 1.15).
гексокіназа- фермент, що каталізує фосфорилювання глюкози за допомогою АТФ. Активний центр ферменту розташовується в щілині між двома доменами. При зв'язуванні гексокінази з глюкозою навколишні її домени змикаються і субстрат виявляється в «пастці», де і відбувається фосфорилювання (див. Рис. 1.15).
 Мал. 1.15. Зв'язування доменів гексокінази з глюкозою
Мал. 1.15. Зв'язування доменів гексокінази з глюкозою
У деяких білках домени виконують самостійні функції, зв'язуючись з різними лігандами. Такі білки називаються багатофункціональними.
3. Ліки - ліганди, що впливають на функцію білків.Взаємодія білків з лігандами специфічно. Однак завдяки конформаційної лабільності білка і його активного центру можна підібрати іншу речовину, яке також могло б взаємодіяти з білком в активному центрі або іншій ділянці молекули.
Речовина, за структурою схоже на природний ліганд, називають структурним аналогом лигандаабо неприродним лигандом. Воно також взаємодіє з білком в активному центрі. Структурний аналог лиганда може як посилювати функцію білка (Агоніст),так і знижувати її (Антагоніст).Ліганд і його структурні аналоги конкурують один з одним за зв'язування з білком в одному центрі. Такі речовини називаються конкурентними модуляторами(Регуляторами) білкових функцій. Багато лікарські препарати діють як інгібітори білків. Деякі з них отримують хімічною модифікацією природних лігандів. Інгібітори білкових функцій можуть бути ліками і отрутами.
Атропін - конкурентний інгібітор М-холінорецепторів.Ацетилхолін - нейромедіатор передачі нервового імпульсу через холінергічні синапси. Для проведення збудження виділився в синаптичну щілину ацетилхолін повинен взаємодіяти з білком - рецептором постсинаптичної мембрани. Виявлено два типи холинорецепторов:
М-рецептор,крім ацетилхоліну вибірково взаємодіє з мускарином (токсином мухомора). М - холінорецептори є на гладких м'язах і при взаємодії з ацетилхоліном викликають їх скорочення;
Н-рецептор,специфічно зв'язується з нікотином. Н-холінорецептори виявлені в синапсах поперечносмугастих скелетних м'язів.
специфічним інгібітором М-холінорецепторівє атропін. Він міститься в рослинах степові і блекоті.
 Атропін має в структурі схожі з ацетилхоліном функціональні групи і їх просторове розташування, тому відноситься до конкурентних інгібіторів М-холінорецепторів. З огляду на, що зв'язування ацетилхоліну з М-холинорецепторами викликає скорочення гладких м'язів, атропін використовують як ліки, що знімає їх спазм (Спазмолітик).Так, відомо застосування атропіну для розслаблення очних м'язів при перегляді очного дна, а також для зняття спазмів при шлунково-кишкових кольках. М-холінорецептори є і в центральній нервовій системі (ЦНС), тому великі дози атропіну можуть викликати небажану реакцію з боку центральної нервової системи: рухове і психічне збудження, галюцинації, судоми.
Атропін має в структурі схожі з ацетилхоліном функціональні групи і їх просторове розташування, тому відноситься до конкурентних інгібіторів М-холінорецепторів. З огляду на, що зв'язування ацетилхоліну з М-холинорецепторами викликає скорочення гладких м'язів, атропін використовують як ліки, що знімає їх спазм (Спазмолітик).Так, відомо застосування атропіну для розслаблення очних м'язів при перегляді очного дна, а також для зняття спазмів при шлунково-кишкових кольках. М-холінорецептори є і в центральній нервовій системі (ЦНС), тому великі дози атропіну можуть викликати небажану реакцію з боку центральної нервової системи: рухове і психічне збудження, галюцинації, судоми.
Дітілін - конкурентний агонист Н-холінорецепторів, що інгібує функцію нервово-м'язових синапсів.
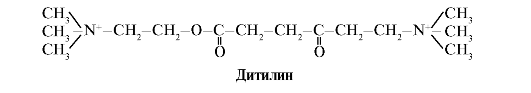 Нервово-м'язові синапси скелетних м'язів містять Н-холінорецептори. Їх взаємодія з ацетилхоліном призводить до м'язових скорочень. При деяких хірургічних операціях, а також в ендоскопічних дослідженнях використовують препарати, що викликають розслаблення скелетних м'язів (Міорелаксанти).До них відноситься дитилин, який є структурним аналогом ацетилхоліну. Він приєднується до Н-холінорецепторів, але на відміну від ацетилхоліну дуже повільно руйнується ферментом - ацетилхолінестеразою. В результаті тривалого відкриття іонних каналів і стійкої деполяризації мембрани порушується проведення нервового імпульсу і відбувається м'язове розслаблення. Спочатку ці властивості були виявлені у отрути кураре, тому такі препарати називають курареподібними.
Нервово-м'язові синапси скелетних м'язів містять Н-холінорецептори. Їх взаємодія з ацетилхоліном призводить до м'язових скорочень. При деяких хірургічних операціях, а також в ендоскопічних дослідженнях використовують препарати, що викликають розслаблення скелетних м'язів (Міорелаксанти).До них відноситься дитилин, який є структурним аналогом ацетилхоліну. Він приєднується до Н-холінорецепторів, але на відміну від ацетилхоліну дуже повільно руйнується ферментом - ацетилхолінестеразою. В результаті тривалого відкриття іонних каналів і стійкої деполяризації мембрани порушується проведення нервового імпульсу і відбувається м'язове розслаблення. Спочатку ці властивості були виявлені у отрути кураре, тому такі препарати називають курареподібними.
ТЕМА 1.3. Денатурація І МОЖЛИВІСТЬ ЇХ СПОНТАННОЇ РЕНАТІВАЦІІ
1. Так як нативна конформація білків підтримується за рахунок слабких взаємодій, зміна складу і властивостей навколишнього білок середовища, вплив хімічних реагентів і фізичних факторів викликають зміну їх конформації (властивість конформационной лабільності). Розрив великої кількості зв'язків призводить до руйнування нативної конформації і денатурації білків.
денатурація білків- це руйнування їх нативної конформації під дією денатуруючих агентів, викликане розривом слабких зв'язків, що стабілізують просторову структуру білка. Денатурація супроводжується руйнуванням унікальною тривимірної структури і активного центру білка і втратою його біологічної активності (рис. 1.16).
Все денатуровані молекули одного білка набувають випадкову конформацию, що відрізняється від інших молекул того ж білка. Радикали амінокислот, що формують активний центр, виявляються просторово віддаленими одна від одної, тобто руйнується специфічний центр зв'язування білка з лігандом. При денатурації первинна структура білків залишається незмінною.
Застосування денатуруючих агентів в біологічних дослідженнях і медицині.У біохімічних дослідженнях перед визначенням в біологічному матеріалі низькомолекулярних сполук зазвичай з розчину спочатку видаляють білки. Для цієї мети найчастіше використовують трихлоруксусную кислоту (ТХУ). Після додавання ТХУ в розчин денатуровані білки випадають в осад і легко видаляються фільтруванням (табл. 1.1.)
У медицині денатурирующие агенти часто застосовують для стерилізації медичного інструменту і матеріалу в автоклавах (денатурує агент - висока температура) і в якості антисептиків (спирт, фенол, хлорамін) для обробки забруднених поверхонь, що містять патогенну мікрофлору.
2. Спонтанна ренатівація білків- доказ детермінованості первинної структури, конформації і функції білків. Індивідуальні білки - це продукти одного гена, які мають ідентичну амінокислотну послідовність і в клітці набувають однакову конформацію. Фундаментальний висновок про те, що в первинній структурі білка вже закладена інформація про його конформації і функції, був зроблений на основі здатності деяких білків (зокрема, рибонуклеази і міоглобіну) до спонтанної ренатіваціі - відновлення їх нативної конформації після денатурації.
Формування просторових структур білка здійснюється способом самозборки - самовільного процесу, при якому поліпептидний ланцюг, що має унікальну первинну структуру, прагне взяти в розчині конформацию з найменшою вільною енергією. Здатність до ренатіваціі білків, що зберігають після денатурації первинну структуру, описана в досвіді з ферментом рібонуклеазою.
Рибонуклеаза - фермент, що руйнує зв'язки між окремими нуклеотидами в молекулі РНК. Цей глобулярний білок має одну поліпептидний ланцюг, третинна структура якої стабілізована безліччю слабких і чотирма дисульфідними зв'язками.
Обробка рибонуклеази сечовиною, яка руйнує водневі зв'язку в молекулі, і відновником, що розривають дисульфідні зв'язки, призводить до денатурації ферменту і втрати його активності.
Видалення денатуруючих агентів діалізом призводить до відновлення конформації і функції білка, тобто до ренатіваціі. (Рис. 1.17).
 Мал. 1.17. Денатурація і ренатівація рибонуклеази
Мал. 1.17. Денатурація і ренатівація рибонуклеази
А - нативная конформація рибонуклеази, в третинної структурі якої є чотири дисульфідні зв'язку; Б - денатурована молекула рибонуклеази;
В - ренатівірованная молекула рибонуклеази з відновленої структурою і функцією
1. Заповніть таблицю 1.2.
Таблиця 1.2. Класифікація амінокислот за полярністю радикалів
2. Напишіть формулу тетрапептіда:
Асп - Про - Фен - Ліз
а) виділіть в пептиді повторювані групи, що утворюють пептидний кістяк, і варіабельні групи, представлені радикалами амінокислот;
б) позначте N- і С-кінці;
в) підкресліть пептидні зв'язку;
г) написати ще один пептид, що складається з тих же амінокислот;
д) підрахуйте кількість можливих варіантів тетрапептіда з аналогічним амінокислотним складом.
3. Поясніть роль первинної структури білків на прикладі порівняльного аналізу двох схожих за структурою і еволюційно близьких пептидних гормонів нейрогипофиза ссавців - окситоцину і вазопресину (табл. 1.3).
Таблиця 1.3. Структура і функції окситоцину і вазопресину
 Для цього:
Для цього:
а) порівняйте склад і послідовність амінокислот двох пептидів;
б) знайдіть схожість первинної структури двох пептидів і схожість їх біологічної дії;
в) знайдіть відмінності в структурі двох пептидів і відмінність їх функцій;
г) зробіть висновок про вплив первинної структури пептидів на їх функції.
4. Опишіть основні етапи формування конформації глобулярних білків (вторинна, третинна структури, поняття про супервторічной структурі). Вкажіть типи зв'язків, що беруть участь у формуванні структур білка. Радикали яких амінокислот можуть брати участь в утворенні гідрофобних взаємодій, іонних, водневих зв'язках.
Наведіть приклади.
5. Дайте визначення поняттю «конформационная лабільність білків», вкажіть причини її існування і значення.
6. Розкрийте сенс наступної фрази: «В основі функціонування білків лежить їх специфічна взаємодія з лігандом», використовуючи терміни і пояснюючи їх значення: конформація білка, активний центр, ліганд, компліментарність, функція білка.
7. На одному з прикладів поясніть, що таке домени і яка їхня роль у функціонуванні білків.
ЗАВДАННЯ ДЛЯ САМОКОНТРОЛЮ
1. Встановіть відповідність.
Функціональна група в радикал амінокислоти:
А. Карбоксильная група Б. Гідроксильна група В гуанідинового група Г. тіольний група Д. Аміногрупа
2. Виберіть правильні відповіді.
Амінокислоти з полярними незарядженими радикалами - це:
A. Цис Б. АСН
B. Гли Г. Три
3. Виберіть правильні відповіді.
Радикали амінокислот:
A. Забезпечують специфічність первинної структури Б. Беруть участь у формуванні третинної структури
B. Розташовуючись на поверхні білка, впливають на його розчинність Г. Формують активний центр
Д. Беруть участь в утворенні пептидних зв'язків
4. Виберіть правильні відповіді.
Гідрофобні взаємодії можуть утворюватися між радикалами амінокислот:
A. Тре Лей Б. Про Три
B. Мет Ілі Г. Тир Ала Д. Вал Фен
5. Виберіть правильні відповіді.
Іонні зв'язку можуть утворюватися між радикалами амінокислот:
A. Глн Асп Б. Apr Ліз
B. Ліз Гли Г. Гіс Асп Д. АСН Apr
6. Виберіть правильні відповіді.
водневі зв'язку можуть утворюватися між радикалами амінокислот:
A. Сер Глн Б. Цис Тре
B. Асп Ліз Г. Гли Асп Д. АСН Тре
7. Встановіть відповідність.
Тип зв'язку, який бере участь у формуванні структури білка:
A. Первинна структура Б. Вторинна структура
B. Третинна структура
Г. Супервторічная структура Д. Конформація.
1. Водневі зв'язку між атомами пептидного остова
2. Слабкі зв'язку між функціональними групами радикалів амінокислот
3. Зв'язки між α-аміно і α-карбоксильними групами амінокислот
8. Виберіть правильні відповіді. трипсин:
A. Протеолітичний фермент Б. Містить два домена
B. гидролизируется крохмаль
Г. Активний центр розташований між доменами. Д. Складається з двох поліпептидних ланцюгів.
9. Виберіть правильні відповіді. атропін:
A. нейромедіаторів
Б. Структурний аналог ацетилхоліну
B. Взаємодіє з Н-холинорецепторами
Г. Підсилює проведення нервового імпульсу через холінергічні синапси
Д. Конкурентний інгібітор М-холінорецепторів
10. Виберіть правильні твердження. У білках:
A. Первинна структура містить інформацію про будову його активного центру
Б. Активний центр формується на рівні первинної структури
B. Конформація жорстко фіксована ковалентними зв'язками
Г. Активний центр може взаємодіяти з групою схожих лигандов
завдяки конформаційної лабільності білків Д. Зміна довкілля, Може впливати на спорідненість активного
центру до ліганду
1. 1-В, 2-Г, 3-Б.
3. А, Б, В, Г.
7. 1-Б, 2-Д, 3-А.
8. А, Б, В, Г.
ОСНОВНІ ТЕРМІНИ І ПОНЯТТЯ
1. Білок, поліпептид, амінокислоти
2. Первинна, вторинна, третинна структури білка
3. Конформація, нативная конформація білка
4. Ковалентні і слабкі зв'язки в білку
5. Конформаційна лабільність
6. Активний центр білка
7. Ліганди
8. згортання білків
9. Структурні аналоги лигандов
10. Доменні білки
11. Прості і складні білки
12. Денатурація білка, денатурирующие агенти
13. Ренатівація білків
Вирішіть завдання
«Структурна організація білків і основи їх функціонування»
1. Основна функція білка - гемоглобіну А (НВА) - транспорт кисню до тканин. У популяції людей відомі множинні форми цього білка із зміненими властивостями і функцією - так звані аномальні гемоглобіни. Наприклад, встановлено, що гемоглобін S, виявлений в еритроцитах хворих серповидно-клітинною анемією (HbS), має низьку розчинність в умовах низького парціального тиску кисню (як це має місце в венозної крові). Це призводить до утворення агрегатів даного білка. Білок втрачає свою функцію, випадає в осад, а еритроцити набувають неправильну форму (Деякі з них утворюють форму серпа) і швидше звичайного руйнуються в селезінці. В результаті розвивається серповидноклітинна анемія.
Єдина відмінність в первинній структурі НВА і виявлено в N-кінцевій ділянці β-ланцюга гемоглобіну. Порівняйте N-кінцеві ділянки β-ланцюга і покажіть, як зміни в первинну структуру білка впливають на його властивості та функції.
 Для цього:
Для цього:
а) напишіть формули амінокислот, за якими розрізняються НВА і порівняйте властивості цих амінокислот (полярність, заряд).
б) зробіть висновок про причини зниження розчинності і порушення транспорту кисню до тканин.
2. На малюнку представлена \u200b\u200bсхема будови білка, що має центр зв'язування з лігандом (активний центр). Поясніть, чому білок має вибірковість у виборі ліганду. Для цього:
а) згадайте, що таке активний центр білка, і розгляньте будову активного центру білка, представленого на малюнку;
б) напишіть формули радикалів амінокислот, що входять до складу активного центру;
в) намалюйте ліганд, який міг би специфічно взаємодіяти з активним центром білка. Вкажіть на ньому функціональні групи, здатні утворити зв'язку з радикалами амінокислот, що входять до складу активного центру;
г) вкажіть типи зв'язків, що виникають між лігандом і радикалами амінокислот активного центру;
д) поясніть, на чому заснована специфічність взаємодії білка з лігандом.
 3.
На малюнку представлений активний центр білка і кілька лігандів.
3.
На малюнку представлений активний центр білка і кілька лігандів.
Визначте, який з лігандів з найбільшою ймовірністю буде взаємодіяти з активним центром білка і чому.
 Які типи зв'язків виникають в процесі утворення комплексу білок-ліганд »?
Які типи зв'язків виникають в процесі утворення комплексу білок-ліганд »?
4. Структурні аналоги природних лігандів білків можуть використовуватися в якості лікарських препаратів для зміни активності білків.
Ацетилхолін - медіатор передачі збудження в нервово-м'язових синапсах. При взаємодії ацетилхоліну з білками - рецепторами постсинаптичної мембрани скелетних м'язів відбувається відкриття іонних каналів і м'язове скорочення. Дітілін - ліки, що застосовується при деяких операціях для розслаблення м'язів, так як він порушує передачу нервового імпульсу через нервово-м'язові синапси. Поясніть механізм дії дитилина як миорелаксирующего препарату. Для цього:
а) напишіть формули ацетилхоліну і дитилина і порівняйте їх структури;
б) опишіть механізм розслабляючу дію дитилина.
5. При деяких захворюваннях у хворого підвищується температура тіла, що розглядають як захисну реакцію організму. Однак високі температури згубні для білків організму. Поясніть, чому при температурі вище 40 ° С порушується функція білків і виникає загроза для життя людини. Для цього згадайте:
1) Будова білків і зв'язку, які утримують його структуру в нативної конформації;
2) Як змінюється структура і функція білків при підвищенні температури ?;
3) Що таке гомеостаз і чому він важливий для підтримки здоров'я людини.
Модульна одиниця 2 олігомерних БЕЛКИ ЯК МІШЕНІ РЕГУЛЯТОРНИХ ВПЛИВІВ. СТРУКТУРНО-ФУНКЦІОНАЛЬНИЙ МНОГООБРАЗИЕ БІЛКІВ. МЕТОДИ РОЗПОДІЛУ І ОЧИЩЕННЯ БІЛКІВ
Цілі вивчення Вміти:
1. Використовувати знання про особливості структури і функцій олігомерних білків для розуміння адаптивних механізмів регуляції їх функцій.
2. Пояснювати роль шаперонов в синтезі і підтримці конформації білків в умовах клітини.
3. Пояснювати різноманіття прояви життя різноманіттям структур і функцій синтезуються в організмі білків.
4. Аналізувати зв'язок структури білків з їх функцією на прикладах порівняння споріднених гемопротеинов - міоглобіну і гемоглобіну, а також представників п'яти класів білків сімейства імуноглобулінів.
5. Застосовувати знання про особливості фізико-хімічних властивостей білків для вибору методів їх очищення від інших білків і домішок.
6. Інтерпретувати результати кількісного та якісного складу білків плазми крові для підтвердження або уточнення клінічного діагнозу.
знати:
1. Особливості будови олігомерних білків і адаптивні механізми регуляції їх функцій на прикладі гемоглобіну.
2. Будова і функції шаперонов і їх значення для підтримки нативної конформації білків в умовах клітини.
3. Принципи об'єднання білків в сімейства по схожості їх конформації і функцій на прикладі імуноглобулінів.
4. Методи поділу білків, засновані на особливостях їх фізико-хімічних властивостей.
5. Електрофорез плазми крові як метод оцінки якісного і кількісного складу білків.
ТЕМА 1.4. ОСОБЛИВОСТІ БУДОВИ І ФУНКЦІОНУВАННЯ олігомерного БІЛКІВ НА ПРИКЛАДІ ГЕМОГЛОБІНУ
1. Багато білки мають в своєму складі кілька поліпептидних ланцюгів. Такі білки називають олігомерними,а окремі ланцюги - протомеров.Протомери в олігомерних білках з'єднані безліччю слабких нековалентних зв'язків (гідрофобних, іонних, водневих). взаємодія
протомеров здійснюється завдяки комплементарностіїх контактуючих поверхонь.
Кількість протомеров в олігомерних білках може сильно варіювати: гемоглобін містить 4 протомеров, фермент аспартатамінотрансфераза - 12 протомеров, а в білок вірусу тютюнової мозаїки входить 2120 протомеров, з'єднаних нековалентними зв'язками. Отже, олігомерні білки можуть мати дуже велику молекулярну масу.
Взаємодія одного протомеров з іншими можна розглядати як окремий випадок взаємодії білка з лігандом, так як кожен протомеров служить лігандом для інших протомеров. Кількість і спосіб з'єднання протомеров в білку називається четвертичной структурою білка.
До складу білків можуть входити однакові або різні за будовою протомери, наприклад, гомодимера - білки, що містять два однакових протомеров, а гетеродімери - білки, що містять два різних протомеров.
Якщо до складу білків входять різні протомери, то на них можуть формуватися відрізняються за структурою центри зв'язування з різними лігандами. При зв'язуванні ліганда з активним центром проявляється функція даного білка. Центр, розташований на іншому протомеров, називається аллостерическим (іншим, відмінним від активного). зв'язуючись з аллостерическим лигандом або ефектором,він виконує регуляторну функцію (рис. 1.18). Взаємодія аллостеріческого центру з ефектором викликає конформаційні зміни в структурі всього олігомерного білка завдяки його конформационной лабільності. Це впливає на спорідненість активного центру до специфічного ліганду і регулює функцію даного білка. Зміна конформації і функції всіх протомеров при взаємодії олігомерного білка хоча б з одним лігандом носить назву кооперативних змін конформації. Ефектори, які посилюють функцію білка, називаються активаторами,а Ефектори, які пригнічують його функцію, - інгібіторами.
Таким чином, у олігомерних білків, а також білків, що мають доменне будова, з'являється нове в порівнянні з мономірними білками властивість - здатність до аллостерічеськой регуляції функцій (регуляції приєднанням до білка різних лігандів). Це можна простежити, порівнюючи структури і функції двох близько споріднених складних білків міоглобіну і гемоглобіну.
 Мал. 1.18. Схема будови дімерная білка
Мал. 1.18. Схема будови дімерная білка
2. Формування просторових структур і функціонування міоглобіну.
Міоглобін (Мв) - білок, що знаходиться в червоних м'язах, основна функція якого - створення запасів О2, необхідних при інтенсивній м'язовій роботі. Мв - складний білок, що містить білкову частина - апоМв і небілкову частина - гем. Первинна структура апоМв визначає його компактну глобулярную конформацию і структуру активного центру, до якого приєднується небілкова частина міоглобіну - гем. Кисень, який надходить з крові в м'язи, зв'язується з Fe + 2 гема в складі міоглобіну. Мв - одновимірний білок, який має дуже високу спорідненість до О2, тому віддача кисню миоглобином відбувається тільки при інтенсивній м'язовій роботі, коли парціальний тиск O 2 різко знижується.
Формування конформації Мв.У червоних м'язах на рибосомах в ході трансляції йде синтез первинної структури Мв, представленої специфічної послідовністю 153 амінокислотних залишків. Вторинна структура Мв містить вісім α-спіралей, званих латинськими літерами від А до Н, між якими є неспіралізованние ділянки. Третинна структура Мв має вигляд компактної глобули, в поглибленні якої між F і Е α-спіралями розташований активний центр (рис. 1.19).
 Мал. 1.19. структура міоглобіну
Мал. 1.19. структура міоглобіну
3. Особливості будови і функціонування активного центру Мв.Активний центр Мв сформований переважно гідрофобними радикалами амінокислот, далеко відстають один від одного в первинній структурі (наприклад, Три 3 9 і Фен 138) До активного центру приєднується погано розчинні у воді ліганди - гем і О2. Гем - специфічний ліганд апоМв (рис. 1.20), основу якого складають чотири піррольних кільця, з'єднаних метенільнимі містками; в центрі розташований атом Fe + 2, з'єднаний з атомами азоту піррольних кілець чотирма координаційними зв'язками. В активному центрі Мв крім гідрофобних радикалів амінокислот є також залишки двох амінокислот з гідрофільними радикалами - Гіс Е 7(Гіс 64) і Гіс F 8(Гіс 93) (рис. 1.21).
 Мал. 1.20. Будова гема - небілкової частини міоглобіну і гемоглобіну
Мал. 1.20. Будова гема - небілкової частини міоглобіну і гемоглобіну
 Мал. 1.21. Розташування гема і O 2 в активному центрі апоміоглобіна і протомеров гемоглобіну
Мал. 1.21. Розташування гема і O 2 в активному центрі апоміоглобіна і протомеров гемоглобіну
Гем через атом заліза ковалентно пов'язаний з Гіс F 8. O 2 приєднується до заліза з іншого боку площині гема. Гіс Е 7 необхідний для правильної орієнтації О2 і полегшує приєднання кисню до Fe + 2 гема
Гіс F 8утворює координаційну зв'язок з Fe + 2 і міцно фіксує гем в активному центрі. Гіс Е 7необхідний для правильної орієнтації в активному центрі іншого ліганда - O 2 при його взаємодії з Fe + 2 гема. Мікрооточення гема створює умови для міцного, але оборотного зв'язування O 2 з Fe +2 і перешкоджає попаданню в гідрофобний активний центр води, що може призвести до його окислення в Fе + 3.
Мономерна будова Мв і його активного центру визначає високу спорідненість білка до О2.
4. олігомерного будова Нв і регуляція спорідненості Нв до О2 лигандами. гемоглобіни людини- сімейство білків, так само як і міоглобін відносяться до складних білок (гемопротеинов). Вони мають тетрамерную будова і містять дві α-ланцюга, але розрізняються за будовою двох інших поліпептидних ланцюгів (2α-, 2х-ланцюга). Будова другий поліпептидного ланцюга визначає особливості функціонування цих форм Нв. Близько 98% гемоглобіну еритроцитів дорослої людини становить гемоглобін А(2α-, 2р-ланцюга).
У період внутрішньоутробного розвитку функціонують два основних типи гемоглобинов: ембріональний Нв(2α, 2ε), який виявляється на ранніх етапах розвитку плода, і гемоглобін F (фетальний)- (2α, 2γ), який приходить на зміну раннього гемоглобіну плода на шостому місяці внутрішньоутробного розвитку і тільки після народження заміщається на Нв А.
Нв А - білок, споріднений міоглобіну (Мв), міститься в еритроцитах дорослої людини. Будова його окремих протомеров аналогічно такому в міоглобіну. Вторинна і третинна структури міоглобіну і протомеров гемоглобіну дуже схожі, незважаючи на те що в первинній структурі їх поліпептидних ланцюгів ідентичні тільки 24 амінокислотних залишку (вторинна структура протомеров гемоглобіну, так само як міоглобін, містить вісім α-спіралей, що позначаються латинськими літерами від А до Н , а третинна структура має вигляд компактної глобули). Але на відміну від міоглобіну гемоглобін має олігомерного будова, складається з чотирьох поліпептидних ланцюгів, з'єднаних нековалентними зв'язками (рис 1.22).
Кожен протомеров Нв пов'язаний з небелковой частиною - гемом і сусідніми протомеров. З'єднання білкової частини Нв з гемом аналогічно такому в міоглобіну: в активному центрі білка гідрофобні частини гема оточені гідрофобними радикалами амінокислот за винятком Гіс F 8 і Гіс Е 7, які розташовані по обидва боки від площини гема і грають аналогічну роль у функціонуванні білка і зв'язуванні його з киснем (див. будова міоглобіну).
 Мал. 1.22. Олігомерного структура гемоглобіну
Мал. 1.22. Олігомерного структура гемоглобіну
Крім того, Гіс Е 7виконує важливу додаткову рольу функціонуванні Нв. Вільний гем має в 25 000 разів вищу спорідненість до СО, ніж до О2. СО в невеликих кількостях утворюється в організмі і, з огляду на його високу спорідненість до гему, він міг би порушувати транспорт необхідного для життя клітин О2. Однак в складі гемоглобіну спорідненість гема до оксиду вуглецю перевищує спорідненість до О2 всього в 200 раз завдяки наявності в активному центрі Гіс Е 7. Залишок цієї амінокислоти створює оптимальні умови для зв'язування гема з O 2 і послаблює взаємодію гема з СО.
5. Основна функція Нв - транспорт О2 з легких в тканини.На відміну від мономерного міоглобіну, що має дуже високу спорідненість до О2 і виконує функцію запасання кисню в червоних м'язах, олігомерного структура гемоглобіну забезпечує:
1) швидке насичення Нв киснем в легенях;
2) здатність Нв віддавати кисень в тканинах при відносно високому парціальному тиску O 2 (20-40 мм рт. Ст.);
3) можливість регуляції спорідненості Нв до О2.
6. Кооперативні зміни конформації протомеров гемоглобіну прискорюють зв'язування O 2 в легенях і віддачу його в тканини. У легких високий парціальний тиск O 2 сприяє зв'язуванню його з Нв в активному центрі чотирьох протомеров (2α і \u200b\u200b2β). Активний центр кожного протомеров, так само як і в миоглобине, розташований між двома α-спіралями (F і Е) в гідрофобному кишені. Він містить небілкову частина - гем, прикріплений до білкової частини безліччю слабких гідрофобних взаємодій і однієї міцнішим зв'язком між Fe 2 + гема і Гіс F 8 (див. Рис. 1.21).
У дезоксігемоглобіне, завдяки цьому зв'язку з Гіс F 8, атом Fe 2 + виступає з площини гема у напрямку до гістидину. Зв'язування O 2 з Fe 2 + відбувається по інший бік гема в області Гіс Е 7 за допомогою єдиної вільної координаційного зв'язку. Гіс Е 7 забезпечує оптимальні умови для зв'язування O 2 з залізом гема.
Приєднання O 2 до атому Fe +2 одного протомеров викликає його переміщення в площину гема, а за ним і залишку гістидину, пов'язаного з ним
 Мал. 1.23. Зміна конформації протомеров гемоглобіну при з'єднанні з O 2
Мал. 1.23. Зміна конформації протомеров гемоглобіну при з'єднанні з O 2
Це призводить до зміни конформації всіх поліпептидних ланцюгів за рахунок їх конформаційної лабільності. Зміна конформації інших ланцюгів полегшує їх взаємодію з наступними молекулами О2.
Четверта молекула О2 приєднується до гемоглобіну в 300 разів легше, ніж перша (рис. 1.24).
 Мал. 1.24. Кооперативні зміни конформації протомеров гемоглобіну при його взаємодії з О2
Мал. 1.24. Кооперативні зміни конформації протомеров гемоглобіну при його взаємодії з О2
У тканинах кожна наступна молекула O 2 відщеплюється легше, ніж попередня, також за рахунок кооперативних змін конформації протомеров.
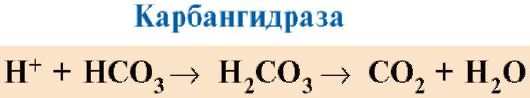
7. CO 2 і Н +, що утворюються при катаболізмі органічних речовин, зменшують спорідненість гемоглобіну до О2 пропорційно їх концентрації. Енергія, необхідна для роботи клітин, виробляється переважно в мітохондріях при окисленні органічних речовин з використанням O 2, що доставляється з легких гемоглобіном. В результаті окислення органічних речовин утворюються кінцеві продукти їх розпаду: СО 2 і K 2 O, кількість яких пропорційно інтенсивності протікають процесів окислення.
СО2 дифузією потрапляє з клітин в кров і проникає в еритроцити, де під дією ферменту карбангідрази перетворюється в вугільну кислоту. Ця слабка кислота дисоціює на протон і бікарбонат іон.
Н + здатні приєднаються до радикалам Гіс 14 6 в α- і β-ланцюгах гемоглобіну, тобто в ділянках, віддалених від гема. Протонирование гемоглобіну знижує його спорідненість до О2, сприяє отщеплению О2 від оксіНв, утворення дезоксіНв і збільшує надходження кисню до тканин пропорційно кількості утворилися протонів (рис. 1.25).
Збільшення кількості звільненого кисню в залежності від збільшення концентрації Н + в еритроцитах називається ефектом Бора (по імені датського фізіолога Християна Бора, що вперше відкрив цей ефект).
У легких високий парціальний тиск кисню сприяє його зв'язування з дезоксіНв, що зменшує спорідненість білка до Н +. Вивільнені протони під дією карбангідрази взаємодіють з бікарбонатами з утворенням СО 2 і Н 2 О
 Мал. 1.25. Залежність спорідненості Нв до О2 від концентрації СО 2 і протонів (ефект Бора):
Мал. 1.25. Залежність спорідненості Нв до О2 від концентрації СО 2 і протонів (ефект Бора):
А- вплив концентрації СО 2 і Н + на вивільнення О2 з комплексу з Нв (ефект Бора); Б- дизельні дезоксигемоглобина в легких, освіту і виділення СО 2.
Утворився СО 2 надходить в альвеолярне простір і видаляється з повітрям, що видихається. Таким чином, кількість вивільняється гемоглобіном кисню в тканинах регулюється продуктами катаболізму органічних речовин: чим інтенсивніше розпад речовин, наприклад при фізичних навантаженнях, тим вище концентрація СО 2 і Н + і тим більше кисню отримують тканини в результаті зменшення спорідненості Нв до О2.
8. аллостеріческого регуляція спорідненості Нв до О2 лигандом - 2,3-біс фосфогліцерат.В еритроцитах з продукту окислення глюкози - 1,3-бісфосфогліцерата синтезується аллостерічеський ліганд гемоглобіну - 2,3-бісфосфогліцерат (2,3-БФГ). У нормальних умовах концентрація 2,3-БФГ висока і її можна порівняти з концентрацією Нв. 2,3-БФГ має сильний негативний заряд -5.
 Бісфосфогліцерат в капілярах тканин, зв'язуючись з дезоксигемоглобином, збільшує вихід кисню в тканини, зменшуючи спорідненість Нв до О2.
Бісфосфогліцерат в капілярах тканин, зв'язуючись з дезоксигемоглобином, збільшує вихід кисню в тканини, зменшуючи спорідненість Нв до О2.
У центрі тетрамерной молекули гемоглобіну знаходиться порожнина. Її утворюють амінокислотні залишки всіх чотирьох протомеров (див. Рис. 1.22). У капілярах тканин протонирование Нв (ефект Бора) призводить до розриву зв'язку між залізом гема і О2. У молекулі
дезоксигемоглобина в порівнянні з оксигемоглобіном виникають додаткові іонні зв'язку, Що з'єднують протомери, внаслідок чого розміри центральної порожнини в порівнянні з оксигемоглобіном збільшуються. Центральна порожнину є місцем приєднання 2,3-БФГ до гемоглобіну. Через відмінності в розмірах центральної порожнини 2,3-БФГ може приєднуватися тільки до дезоксігемоглобіну.
2,3-БФГ взаємодіє з гемоглобіном в ділянці, віддаленому від активних центрів білка і відноситься до аллостерическим(Регуляторним) лигандам, а центральна порожнина Нв є аллостерическим центром.2,3-БФГ має сильний негативний заряд і взаємодіє з п'ятьма позитивно зарядженими групами двох β-ланцюгів Нв: N-кінцевий α-аміногрупою Вал і радикалами Ліз 82 Гіс 143 (рис. 1.26).
 Мал. 1.26. БФГ в центральній порожнини дезоксигемоглобина
Мал. 1.26. БФГ в центральній порожнини дезоксигемоглобина
БФГ зв'язується з трьома позитивно зарядженими групами в кожній β-ланцюга.
У капілярах тканин утворюється дезоксигемоглобін взаємодіє з 2,3-БФГ і між позитивно зарядженими радикалами β-ланцюгів і негативно зарядженим лігандом утворюються іонні зв'язку, які змінюють конформацію білка і знижують спорідненість Нв до О2. Зменшення спорідненості Нв до О2 сприяє більш ефективному виходу О2 в тканини.
У легких при високому парціальному тиску кисень взаємодіє з Нв, приєднуючись до заліза гема; при цьому змінюється конформація білка, зменшується центральна порожнина і відбувається витіснення 2,3-БФГ з аллостеріческого центру
Таким чином, олігомерні білки володіють новими в порівнянні з мономірними білками властивостями. Приєднання лігандів на ділянках,
просторово віддалених один від одного (аллостеріческіх), здатне викликати конформаційні зміни у всій білкової молекулі. Завдяки взаємодії з регуляторними лигандами відбувається зміна конформації і адаптація функції білкової молекули до змін навколишнього середовища.
ТЕМА 1.5. ПІДТРИМКА нативної конформації БІЛКІВ В УМОВАХ клітки
У клітинах в процесі синтезу поліпептидних ланцюгів, їх транспорту через мембрани до відповідних відділів клітини, в процесі фолдинга (формування нативної конформації) і при складанні олігомерних білків, а також у період їх функціонування в структурі білків виникають проміжні, схильні до агрегації, нестабільні конформації. Гідрофобні радикали, в нативної конформації зазвичай заховані всередині білкової молекули, в нестабільній конформації виявляються на поверхні і прагнуть до об'єднання з такими ж погано розкладаються у воді групами інших білків. У клітинах всіх відомих організмів виявлені спеціальні білки, які забезпечують оптимальний фолдінг білків клітини, стабілізують їх нативну конформацію при функціонуванні і, що особливо важливо, підтримують структуру і функції внутрішньоклітинних білків при порушенні гомеостазу. Ці білки отримали назву «Шаперони»,що в перекладі з французької означає «няня».
1. Молекулярні шаперони і їх роль в запобіганні денатурації білків.
Шаперони (Ш) класифікуються за масою субодиниць. Високомолекулярні шаперони мають масу від 60 до 110 кБ. Серед них найбільш вивчені три класи: Ш-60, Ш-70 та Ш-90. Кожен клас включає сімейство споріднених білків. Так, до складу Ш-70 входять білки з молекулярною масою від 66 до 78 кД. Низькомолекулярні шаперони мають молекулярну масу від 40 до 15 кД.
Серед шаперонов розрізняють конститутивнібілки, високий базальний синтез яких не залежить від стресових впливів на клітини організму, і індуцібельная,синтез яких в нормальних умовах йде слабо, але різко зростає при стресових впливах. Індуцібельная шаперони називають також «білками теплового шоку», так як вперше вони були виявлені в клітинах, які піддавалися впливу високих температур. У клітинах через високу концентрацію білків мимовільна ренатівація частково денатурованих білків утруднена. Ш-70 можуть запобігати почався процес денатурації і сприяти відновленню нативної конформації білків. Молекулярні шаперони-70- висококонсерватівний клас білків, що знаходяться у всіх відділах клітини: цитоплазмі, ядрі, ЕПР, мітохондріях. На Карбоксильні кінці єдиною поліпептидного ланцюга Ш-70 є ділянка, який представляє собою борозенку, здатний взаємодіяти з пептидами довжиною
від 7 до 9 амінокислотних залишків, збагачених гідрофобними радикалами. Такі ділянки в глобулярних білках зустрічаються приблизно через кожні 16 амінокислот. Ш-70 здатні захищати білки від температурної інактивації і відновлювати конформацию і активність частково денатурованих білків.
2. Роль шаперонов в згортання білків.При синтезі білків на рибосомі N-кінцева область поліпептиду синтезується раніше С-кінцевий. Для формування нативної конформації необхідна повна амінокислотна послідовність білка. У процесі синтезу білків шаперони-70, завдяки будові їх активного центру, здатні закривати схильні до агрегації ділянки поліпептиду, збагачені гідрофобними радикалами амінокислот до завершення синтезу (рис 1.27, А).
 Мал. 1.27. Участь шаперонов в згортання білків
Мал. 1.27. Участь шаперонов в згортання білків
А - участь шаперонов-70 в запобіганні гідрофобних взаємодій між ділянками синтезирующегося поліпептиду; Б - формування нативної конформації білка в шапероновом комплексі
Багато високомолекулярні білки, які мають складну конформацію, наприклад доменне будова, здійснюють фолдінг в спеціальному просторі, сформованому Ш-60. Ш-60функціонують у вигляді олігомерного комплексу, що складається з 14 субодиниць. Вони формують два порожніх кільця, кожне з яких складається з семи субодиниць, ці кільця з'єднані один з одним. Кожна субодиниця Ш-60 складається з трьох доменів: апикального (верхівкового), збагаченого гідрофобними радикалами, зверненими в порожнину кільця, проміжного і екваторіального (рис. 1.28).
 Мал. 1.28. Структура шаперонінового комплексу, що складається з 14 Ш-60
Мал. 1.28. Структура шаперонінового комплексу, що складається з 14 Ш-60
А - вид збоку; Б - вид зверху
Синтезовані білки, що мають на поверхні елементи, характерні для несвернутих молекул, зокрема гідрофобні радикали, потрапляють в порожнину шаперонових кілець. У специфічному середовищі цих порожнин відбувається перебір можливих конформацій, поки не буде знайдена єдина, енергетично найбільш вигідна (рис. 1.27, Б). Формування конформаций і вивільнення білка супроводжується гідролізом АТФ в екваторіальній області. Зазвичай такий шаперонозавісімий фолдінг вимагає витрат значної кількості енергії.
Крім участі у формуванні тривимірної структури білків і ренатіваціі частково денатурованих білків, шаперони також необхідні для протікання таких фундаментальних процесів, як збірка олігомерних білків, впізнавання і транспорт в лізосоми денатурованих білків, транспорт білків через мембрани, участь в регуляції активності білкових комплексів.
ТЕМА 1.6. МНОГООБРАЗИЕ БІЛКІВ. РОДИНИ БІЛКІВ НА ПРИКЛАДІ імуноглобулінів
1. Білки відіграють вирішальну роль в життєдіяльності окремих клітин і всього багатоклітинного організму, А їх функції дивно різноманітні. Це визначається особливостями первинної структури і конформаций білків, унікальністю будови активного центру і здатністю пов'язувати специфічні ліганди.
Лише дуже невелика частина всіх можливих варіантів пептидних ланцюгів може прийняти стабільну просторову структуру; більшість
з них може приймати безліч конформацій з приблизно однаковою енергією Гіббса, але з різними властивостями. Первинна структура більшості відомих білків, відібраних біологічною еволюцією, забезпечує виняткову стабільність однією з конформаций, яка визначає особливості функціонування цього білка.
2. Сімейства білків.В межах одного біологічного виду заміни амінокислотних залишків можуть призводити до виникнення різних білків, що виконують споріднені функції і мають гомологічні послідовності амінокислот. Такі родинні білки мають разюче схожі конформації: кількість і взаємне розташування α-спіралей і (або) β-структур, більшість поворотів і вигинів поліпептидних ланцюгів схожі або ідентичні. Білки з гомологічними ділянками поліпептидного ланцюга, схожою конформацией і спорідненими функціями виділяють в сімейства білків. Приклади сімейств білків: серинові протеїнази, сімейство імуноглобулінів, сімейство міоглобіну.
серинові протеїнази- сімейство білків, що виконують функцію протеолітичних ферментів. До них відносяться травні ферменти - хімотрипсин, трипсин, еластаза та багато чинників згортання крові. Ці білки мають в 40% положень ідентичні амінокислоти і дуже близьку конформацию (рис. 1.29).

Мал. 1.29. Просторові структури еластази (А) і хімотрипсину (Б)
Деякі амінокислотні заміни привели до зміни субстратної специфічності цих білків і виникнення функціонального різноманіття всередині сімейства.
3. Сімейство імуноглобулінів.В роботі імунної системи величезну роль грають білки суперсімейства імуноглобулінів, яке включає в себе три сімейства білків:
Антитіла (імуноглобуліни);
Рецептори Т-лімфоцитів;
Білки головного комплексу гістосумісності - МНС 1-го і 2-го класів (Major Histocompatibility Complex).
Всі ці білки мають доменне будова, складаються з гомологічних іммуноподобних доменів і виконують подібні функції: взаємодіють з чужорідними структурами, або розчиненими в крові, лімфі або міжклітинної рідини (антитіла), або знаходяться на поверхні клітин (власних або чужорідних).
4. Антитіла- специфічні білки, що виробляються В-лімфоцитами у відповідь на потрапляння в організм чужорідної структури, званої антигеном.
Особливості будови антитіл
Найпростіші молекули антитіл складаються з чотирьох поліпептидних ланцюгів: двох ідентичних легких - L, що містять близько 220 амінокислот, і двох ідентичних важких - Н, що складаються з 440-700 амінокислот. Всі чотири ланцюга в молекулі антитіла з'єднані безліччю нековалентних зв'язків і чотирма дисульфідними зв'язками (рис. 1.30).
Легкі ланцюги антитіла складаються з двох доменів: вариабельного (VL), що знаходиться в N-кінцевій області поліпептидного ланцюга, і константного (CL), розташованого на С-кінці. Важкі ланцюги зазвичай мають чотири домена: один варіабельний (VH), що знаходиться на N-кінці, і три константних (CH1, CH2, СНЗ) (див. Рис. 1.30). Кожен домен імуноглобуліну має β-складчасту суперструктуру, в якій два залишку цистеїну з'єднані дисульфідній зв'язком.
Між двома константними доменами СН1 і СН2 є ділянка, що містить велике число залишків проліну, які перешкоджають формуванню вторинної структури і взаємодії сусідніх Н-ланцюгів на цьому відрізку. Ця шарнірна область надає молекулі антитіла гнучкість. Між варіабельними доменами важких і легких ланцюгів знаходяться два ідентичних антигензв'язуючих ділянки (активні центри для зв'язування антигенів), тому такі антитіла часто називають біваленте.У зв'язуванні антигену з антитілом бере участь не вся амінокислотна послідовність варіабельних ділянок обох ланцюгів, а всього лише 20-30 амінокислот, розташованих в гіперваріабельних областях кожної ланцюга. Саме ці області визначають унікальну здатність кожного виду антитіла взаємодіяти з відповідним комплементарних антигеном.
Антитіла - одна з ліній захисту організму проти внедрившихся чужорідних організмів. Їх функціонування можна розділити на два етапи: перший етап - впізнавання і зв'язування антигену на поверхні чужорідних організмів, що можливо завдяки наявності в структурі антитіла антигензв'язуючих ділянок; другий етап - ініціація процесу інактивації і руйнування антигену. Специфічність другого етапу залежить від класу антитіл. Існує п'ять класів важких ланцюгів, що відрізняються один від одного за будовою константних доменів: α, δ, ε, γ і μ, відповідно до яких розрізняють п'ять класів імуноглобулінів: A, D, Е, G і М.
Особливості будови важких ланцюгів надають шарнірним ділянкам і С-кінцевим областям важких ланцюгів характерну для кожного класу конформацию. Після зв'язування антигену з антитілом конформаційні зміни константних доменів визначають шлях видалення антигену.
 Мал. 1. 30. Доменне будова IgG
Мал. 1. 30. Доменне будова IgG
імуноглобуліни М
Імуноглобуліни М мають дві форми.
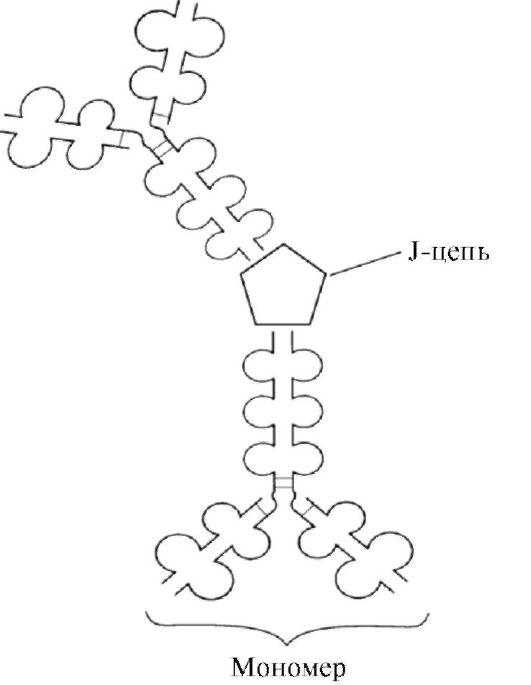
мономерна форма- 1-й клас антитіл, що продукується розвиваються В-лімфоцитів. Згодом багато В-клітини перемикаються на вироблення інших класів антитіл, але з тим же антигензв'язуючих ділянкою. IgM вбудовується в мембрану і виконує роль антігенраспознающіх рецептора. Вбудовування IgM в мембрану клітин можливо завдяки наявності в хвостовій частині ділянки 25 гідрофобних амінокислотних залишків.
Секреторна форма IgMмістить п'ять мономерних субодиниць, пов'язаних один з одним дисульфідними зв'язками і додаткової поліпептидного J-ланцюгом (рис. 1.31). Важкі ланцюги мономерів цієї форми не містять гидрофобной хвостовій частині. Пентамер має 10 центрів зв'язування з антигеном і тому ефективний в розпізнаванні і видаленні вперше потрапив в організм антигену. Секреторна форма IgM - основний клас антитіл, секретується в кров при первинному імунній відповіді. Зв'язування IgM з антигеном змінює конформацію IgM і індукує зв'язування його з першим білковим компонентом системи комплементу (система комплементу - набір білків, що беруть участь у знищенні антигену) і активацію цієї системи. Якщо антиген розташований на поверхні мікроорганізму, система комплементу викликає порушення цілісності клітинної мембрани і загибель бактеріальної клітини.
імуноглобуліни G
У кількісному відношенні цей клас імуноглобулінів переважає в крові (75% від усіх Ig). IgG - мономери, основний клас антитіл, що секретується в кров при вторинному імунній відповіді. Після взаємодії IgG з поверхневими антигенами мікроорганізмів комплекс антиген-антитіло здатний зв'язувати і активувати білки системи комплементу або може взаємодіяти зі специфічними рецепторами макрофагів і нейтрофілів. Взаємодія з фагоцитами призводить
 Мал. 1.31. Будова секреторною форми IgM
Мал. 1.31. Будова секреторною форми IgM
до поглинання комплексів антиген-антитіло і руйнування їх в фагосомах клітин. IgG - єдиний клас антитіл, які здатні проникати через плацентарний бар'єр і забезпечувати внутрішньоутробну захист плода від інфекцій.
імуноглобуліни А
Основний клас антитіл, присутній в секретах (молоці, слині, секрети дихальних шляхів і кишкового тракту). IgA секретируются в основному в димерной формі, де мономери зв'язані один з одним через додаткову J-ланцюг (рис. 1.32).
IgA не взаємодіють з системою комплементу і фагоцитуючими клітинами, але, зв'язуючись з мікроорганізмами, антитіла перешкоджають їх приєднання до епітеліальних клітин і проникненню в організм.
імуноглобуліни Е
Імуноглобуліни Е представлені мономерами, в яких важкі ε-ланцюга містять, так само як і μ-ланцюга імуноглобулінів М, один варіабельний і чотири константних домену. IgE після секреції зв'язуються своїми
 Мал. 1.32. будова IgA
Мал. 1.32. будова IgA
З-кінцевими ділянками з відповідними рецепторами на поверхні огрядних клітин і базофілів. В результаті вони стають рецепторами для антигенів на поверхні цих клітин (рис. 1.33).
 Мал. 1.33. Взаємодія IgE з антигеном на поверхні гладкої клітини
Мал. 1.33. Взаємодія IgE з антигеном на поверхні гладкої клітини
Після того як відбувається приєднання антигену до відповідних антигензв'язуючих ділянкам IgE, клітини отримують сигнал до секреції біологічно активних речовин (Гістаміну, серотоніну), які в великій мірі відповідальні за розвиток запальної реакції і за прояв таких алергічних реакцій, як астма, кропив'янка, сінна лихоманка.
імуноглобуліни D
Імуноглобуліни D виявлені в сироватці в дуже невеликій кількості, вони є мономерами. У важких δ-ланцюгах є один варіабельний і три константних домену. IgD виконують роль рецепторів В-лімфоцитів, інші функції поки невідомі. Взаємодія специфічних антигенів з рецепторами на поверхні В-лімфоцитів (IgD) призводить до передачі цих сигналів в клітку і включення механізмів, що забезпечують розмноження даного клону лімфоцитів.
ТЕМА 1.7. ФІЗИКО-хімічні властивості БІЛКІВ І МЕТОДИ ЇХ РОЗПОДІЛУ
1. Індивідуальні білки розрізняються за фізико-хімічними властивостями:
Формі молекул;
Молекулярної масі;
Сумарному заряду, величина якого залежить від співвідношення аніонних і катіонних груп амінокислот;
Співвідношенню полярних і неполярних радикалів амінокислот на поверхні молекул;
Ступеня стійкості до впливу різних денатуруючих агентів.
2. Розчинність білків залежитьвід властивостей білків, перерахованих вище, а також від складу середовища, в якій розчиняється білок (значення рН, сольового складу, температури, наявності інших органічних речовин, здатних взаємодіяти з білком). Величина заряду білкових молекул - один з факторів, що впливають на їх розчинність. При втраті заряду в ізоелектричної точці білки легше агрегує і випадають в осад. Це особливо характерно для денатурованих білків, у яких на поверхні виявляються гідрофобні радикали амінокислот.
На поверхні білкової молекули є як позитивно, так і негативно заряджені радикали амінокислот. Кількість цих груп, а отже, і сумарний заряд білків залежать від рН середовища, тобто співвідношення концентрації Н + - і ОН --груп. У кислому середовищіпідвищення концентрації Н + призводить до придушення дисоціації карбоксильних груп -СОО - + Н +\u003e - СООН і зниження негативного заряду білків. В лужному середовищі зв'язування надлишку ВІН - протонами, що утворюються при дисоціації аміногруп -NH 3 + + ОН - - NH 2 + Н 2 О з утворенням води, призводить до зменшення позитивного заряду білків. Значення рН, при якому білок має сумарний нульовий заряд, називається ізоелектричної точкою (ВЕТ).У ВЕТ число позитивно і негативно заряджених груп однаково, тобто білок знаходиться в ізоелектричному стані.
3. Поділ індивідуальних білків.Особливості будови і функціонування організму залежать від набору білків, що синтезуються в ньому. Вивчення будови і властивостей білків є неможливим без їх виділення з клітини і очищення від інших білків та органічних молекул. Стадії виділення і очищення індивідуальних білків:
руйнування клітиндосліджуваної тканини і отримання гомогенату.
Поділ гомогената на фракціїцентрифугуванням, отримання ядерної, мітохондріальної, цитозольних чи іншої фракції, що містить шуканий білок.
Виборча теплова денатурація- короткочасне нагрівання розчину білків, при якому можна видалити частину денатурованих білкових домішок (в тому випадку, якщо білок щодо термостабилен).
Висолювання.Різні білки випадають в осад при різних концентраціях солі в розчині. Поступово підвищуючи концентрацію солі, можна отримати ряд окремих фракцій з переважним вмістом виділяється білка в одній з них. Найбільш часто для фракціонування білків використовують сульфат амонію. Білки з найменшою розчинністю випадають в осад при невеликих концентраціях солей.
Гель-фільтрація- метод просіювання молекул через набряклі гранули сефадексе (тривимірні полісахаридні ланцюги декстрану, що мають пори). Швидкість проходження білків через колонку, заповнену сефадексом, буде залежати від їх молекулярної маси: чим менше маса молекул білка, тим легше вони проникають всередину гранул і довше там затримуються, чим більше маса, тим швидше вони елюіруют з колонки.
ультрацентрофугування- метод, який полягає в тому, що білки в центрифужной пробірці поміщають в ротор ультрацентрифуги. При обертанні ротора швидкість осідання білків пропорційна їх молекулярній масі: фракції важчих білків розташовані ближче до дна пробірки, легші - ближче до поверхні.
електрофорез- метод, в основі якого лежать відмінності в швидкості руху білків в електричному полі. Ця величина пропорційна заряду білків. Електрофорез білків проводять на папері (в цьому випадку швидкість руху білків пропорційна тільки їх заряду) або в поліакриламідному гелі з певною величиною пір (швидкість руху білків пропорційна їх заряду і молекулярної масі).
ионообменная хроматографія- метод фракціонування, заснований на зв'язуванні іонізованих груп білків з протилежно зарядженими групами іонообмінних смол (нерозчинних полімерних матеріалів). Міцність зв'язування білка зі смолою пропорційна заряду білка. Білки, адсорбовані на іонообмінному полімері, можна змити розчинами NaCl зі зростаючими концентраціями; чим менше заряд білка, тим менша концентрація NaCl потрібно, щоб змити білок, пов'язаний з йоногенних групами смоли.
аффинная хроматографія- найбільш специфічний метод виділення індивідуальних белков.К інертному полімеру ковалентно приєднується ліганд будь-якого білка. При пропущенні розчину білків через колонку з полімером за рахунок комплементарного зв'язування білка з лігандом на колонці адсорбується тільки специфічний для даного ліганду білок.
діаліз- метод, застосовуваний для видалення низькомолекулярних сполук з розчину виділяється білка. Метод заснований на нездатності білків проходити через напівпроникну мембрану на відміну від низькомолекулярних речовин. Застосовується для очищення білків від низькомолекулярних домішок, наприклад від солей після висолювання.
ЗАВДАННЯ ДЛЯ позааудиторної роботи
1. Заповніть табл. 1.4.
Таблиця 1.4. Порівняльний аналіз структури і функцій родинних білків - міоглобіну і гемоглобіну
а) згадайте будову активного центру Мв і нь. Яку роль відіграють гідрофобні радикали амінокислот у формуванні активних центрів цих білків? Опишіть будову активного центру Мв і нь і механізми приєднання до нього лігандів. Яку роль відіграють залишки Гіс F 8 і Гіс Е 7 в функціонуванні активного центру Мв інв?
б) якими новими властивостями в порівнянні з мономірним миоглобином володіє близько споріднений олігомерного білок - гемоглобін? Поясніть роль кооперативних змін конформації протомеров в молекулі гемоглобіну, вплив концентрацій СО 2 і протонів на спорідненість гемоглобіну до кисню, а також роль 2,3-БФГ в аллостерічеськой регуляції функції нь.
2. Дайте характеристику молекулярним шаперон, звертаючи увагу на зв'язок їх структури з функцією.
3. Які білки об'єднують в сімейства? На прикладі сімейства імуноглобулінів визначте подібні риси будови і родинні функції білків цього сімейства.
4. Часто для біохімічних і медичних цілей потрібні очищені індивідуальні білки. Поясніть, на яких фізико-хімічні властивості білків засновані використовувані методи їх розподілу та очищення.
ЗАВДАННЯ ДЛЯ САМОКОНТРОЛЮ
1. Виберіть правильні відповіді.
Функції гемоглобіну:
A. Транспорт О2 з легких в тканини Б. Транспорт Н + з тканин в легені
B. Підтримання сталості рН крові Г. Транспорт СО 2 з легких в тканини
Д. Транспорт СО 2 з тканин в легені
2. Виберіть правильні відповіді. лігандомα -протомера Нв є:A. Гем
Б. Кисень
B. СО Г. 2,3-БФГ
Д. β-протомеров
3. Виберіть правильні відповіді.
Гемоглобін на відміну від міоглобіну:
A. Має четвертинних структуру
Б. Вторинна структура представлена \u200b\u200bтільки α-спіралями
B. Відноситься до складних білок
Г. Взаємодіє з аллостерическим лигандом Д. ковалентних пов'язаний з гемом
4. Виберіть правильні відповіді.
Спорідненість Нв до О2 зменшується:
A. При приєднанні однієї молекули О2 Б. При відщепленні однієї молекули О2
B. При взаємодії з 2,3-БФГ
Г. При приєднанні до протомеров Н + Д. При зниженні концентрації 2,3-БФГ
5. Встановіть відповідність.
Для типів Нв характерно:
A. В дезокси формі утворює фібрилярні агрегати Б. Містить в складі дві α- і дві δ-ланцюга
B. Переважна форма Нв в еритроцитах дорослої людини Г. В активному центрі містить гем з Fе + 3
Д. Містить в складі дві α- і дві γ-ланцюга 1. НВА 2.
6. Встановіть відповідність.
Ліганди Нв:
A. Зв'язується з Нв в аллостеріческому центрі
Б. Має дуже високу спорідненість до активного центру Нв
B. Приєднуючись, підвищує спорідненість Нв до O 2 Г. Окисляє Fe + 2 в Fе + 3
Д. Утворює ковалентний зв'язок з гісF8
7. Виберіть правильні відповіді.
шаперони:
A. Білки, присутні у всіх відділах клітини
Б. Синтез посилюється при стресових впливах
B. Беруть участь в гідролізі денатурованих білків
Г. Беруть участь в підтримці нативной конформації білків
Д. Створюють органели, в яких формується конформація білків
8. Установіть відповідність. імуноглобуліни:
A. Секреторна форма має пентамерную форму
Б. Клас Ig, що проникають через плацентарний бар'єр
B. Ig - рецептор огрядних клітин
Г. Основний клас Ig, присутніх в секретах епітеліальних клітин. Д. Рецептор В-лімфоцитів, активація якого забезпечує розмноження клітин
9. Виберіть правильні відповіді.
Імуноглобуліни Е:
A. Виробляються макрофагами Б. Мають важкі ε-ланцюга.
B. Вбудовуються в мембрану Т-лімфоцитів
Г. Виконують роль мембранних рецепторів антигенів на огрядних клітках і базофілів
Д. Відповідальні за прояв алергічних реакцій
10. Виберіть правильні відповіді.
Метод поділу білків заснований на відмінностях в їх молекулярній масі:
A. Гель-фільтрація
Б. Ультрацентрофугування
B. Електрофорез в поліакриламідному гелі Г. Ионообменная хроматографія
Д. Аффинная хроматографія
11. Виберіть правильну відповідь.
Метод поділу білків заснований на відмінностях в їх розчинності в воді:
A. Гель-фільтрація Б. Висолювання
B. Ионообменная хроматографія Г. Аффинная хроматографія
Д. Електрофорез в поліакриламідному гелі
ЕТАЛОНИ ВІДПОВІДЕЙ До «ЗАВДАНЬ ДЛЯ САМОКОНТРОЛЮ»
1. А, Б, В, Д
2. А, Б, В, Д
5. 1-В, 2-А, 3-Г
6. 1-В, 2-Б, 3-А
7. А, Б, Г, Д
8. 1-Г; 2-Б, 3-В
ОСНОВНІ ТЕРМІНИ І ПОНЯТТЯ
1. Олігомерні білки, протомеров, четвертичная структура білків
2. Кооперативні зміни конформації протомеров
3. Ефект Бора
4. аллостеріческого регуляція функцій білків, аллостерічеський центр і аллостерічеський ефектор
5. Молекулярні шаперони, білки теплового шоку
6. Сімейства білків (серинові протеази, імуноглобуліни)
7. IgM-, G-, Е, А-зв'язок структури з функцією
8. Сумарний заряд білків, ізоелектричної точка білків
9. Електрофорез
10. Висолювання
11. Гель-фільтрація
12. Ионообменная хроматографія
13. Ультрацентрофугування
14. Аффинная хроматографія
15. Електрофорез білків плазми крові
ЗАВДАННЯ ДЛЯ аудиторної роботи
1. Порівняйте залежності ступенів насичення гемоглобіну (Нв) і міоглобіну (Mb) киснем від його парціального тиску в тканинах
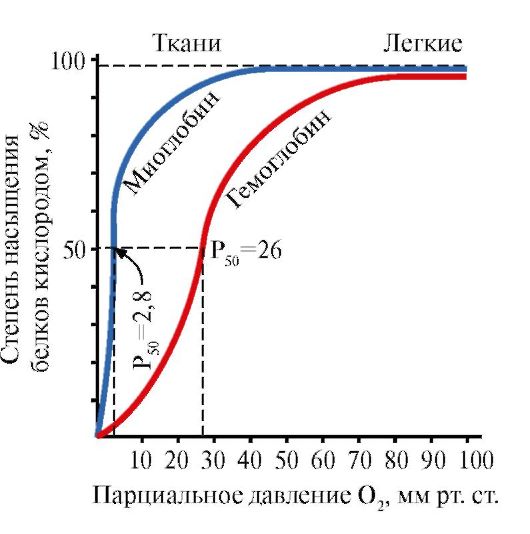 Мал. 1.34. Залежність насичення Мв інькиснем від його парціального тиску
Мал. 1.34. Залежність насичення Мв інькиснем від його парціального тиску
Зверніть увагу, що форма кривих насичення білків киснем різна: для міоглобіну - гіпербола, для гемоглобіну - сігмоідной форма.
1. порівняйте значення парціального тиску кисню, при якому Мв і Нв насичені О2 на 50%. Який з цих білків характеризується більш високою спорідненістю до О2?
2. які особливості будови Мв визначають його високу спорідненість до О2?
3. які особливості будови Нв дозволяють йому віддавати О2 в капілярах покояться тканин (при відносно високому парціальному тиску О2) і різко збільшувати цю віддачу в працюючих м'язах? Яка властивість олігомерних білків забезпечує цей ефект?
4. обчисліть, скільки О2 (в%) віддає оксигенований гемоглобін спочиває і працюючому м'язі?
5. зробіть висновки про зв'язок структури білка з його функцією.
2. Кількість вивільняється гемоглобіном кисню в капілярах залежить від інтенсивності процесів катаболізму в тканинах (ефект Бора). Як зміни метаболізму в тканинах регулюють спорідненість НЬ до O 2? Вплив СО 2 і Н + на спорідненість НЬ до О2
 1. опишіть ефект Бора.
1. опишіть ефект Бора.
2. в якому напрямку протікає процес, представлений на схемі:
а) в капілярах легких;
б) в капілярах тканин?
3. у чому фізіологічне значення ефекту Бора?
4. чому взаємодія Нв з Н + на ділянках, віддалених від гема, змінює спорідненість білка до О2?
3. Спорідненість НЬв до О2 залежить від концентрації його ліганда - 2,3-біфосфо- гліцерата, який є аллостерическим регулятором спорідненості Нв до О2. Чому взаємодія ліганда в ділянці, віддаленому від активного центру, впливає на функцію білка? Як 2,3-БФГ регулює спорідненість НЬ до О2? Для вирішення завдання дайте відповідь на наступні питання:
1. де і з чого синтезується 2.3-біфосфогліцератом (2,3-БФГ)? Напишіть його формулу, вкажіть заряд даної молекули.
2. з якою формою гемоглобіну (окси або дезокси) взаємодіє БФГ і чому? В якій ділянці молекули Нв відбувається взаємодія?
3. в якому напрямку протікає процес, представлений на схемі
а) в капілярах тканин;
б) в капілярах легких?
4. де повинна бути вищою концентрація комплексу
Нв-2,3-БФГ:
а) в капілярах м'язів, що знаходяться в стані спокою,
б) в капілярах працюючих м'язів (за умови однакової концентрації БФГ в еритроцитах)?
5. як зміниться спорідненість Нв до кисню при адаптації людини до умов високогір'я, якщо концентрація БФГ в еритроцитах при цьому підвищується? У чому фізіологічне значення цього явища?
4. Руйнування 2,3-БФГ при зберіганні консервованої крові порушує функції Нв. Як зміниться спорідненість Нв до O 2 в консервованої крові, якщо концентрація 2,3-БФГ в еритроцитах може зменшаться з 8 до 0,5 ммоль / л. Чи можна переливати таку кров тяжкохворим пацієнтам, якщо концентрація 2,3-БФГ відновлюється не раніше ніж через три доби? Чи можна додаванням в кров 2,3-БФГ відновити функції еритроцитів?
5. Згадайте будову найпростіших молекул імуноглобулінів. Яку роль відіграють імуноглобуліни в роботі імунної системи? Чому Ig часто називають біваленте? Як структура Ig пов'язана з їх функцією? (Опишіть на прикладі будь-якого класу імуноглобулінів.)
Фізико-хімічні властивості білків і методи їх розподілу.
6. Як сумарний заряд білка впливає на його розчинність?
а) визначте сумарний заряд пептиду при рН 7
Ала-Гли-Тре-Про-Асп-Ліз-Цис
б) як зміниться заряд цього пептиду при рН\u003e 7, рН<7, рН <<7?
в) що таке ізоелектричної точка білка (ВЕТ) і в якому середовищі лежить
ВЕТ даного пептиду?
г) при якому значенні рН буде спостерігатися найменша розчинність даного пептиду.
7. Чому кисле молоко на відміну від свіжого при кип'ятінні «згортається» (тобто білок молока казеїн випадає в осад)? У свіжому молоці молекули казеїну мають негативний заряд.
8. Для поділу індивідуальних білків використовується метод гельфільтраціі. Суміш, що містить білки А, В, С з молекулярними масами, рівними відповідно 160 000, 80 000 і 60 000, аналізували методом гель-фільтрації (рис. 1.35). Гранули набряклого гелю проникні для білків з молекулярною масою менше 70 000. Який принцип лежить в основі даного методу поділу? Який з графіків правильно відображає результати фракціонування? Вкажіть порядок виходу білків А, В і С з колонки.
 Мал. 1.35. Використання методу гель-фільтрації для розділення білків
Мал. 1.35. Використання методу гель-фільтрації для розділення білків
9. На рис. 1.36, А представлена \u200b\u200bсхема електрофорезу на папері білків сироватки крові здорової людини. Відносні кількості білкових фракцій, отриманих за допомогою цього методу, становлять: альбуміни 54-58%, α 1-глобуліни 6-7%, α 2-глобуліни 8-9%, β-глобуліни 13%, γ-глобуліни 11-12% .
 Мал. 1.36 Електрофорез на папері білків плазми крові здорової людини (А) і хворого (Б)
Мал. 1.36 Електрофорез на папері білків плазми крові здорової людини (А) і хворого (Б)
I - γ-глобуліни; II - β-глобуліни; III -α 2 глобулін; IV -α 2 глобулін; V - альбуміни
Багато хвороб супроводжуються кількісними змінами в складі сироваткових білків (диспротеинемии). Характер цих змін враховується при постановці діагнозу та оцінки тяжкості та стадії захворювання.
Як відомо, білки - основа зародження життя на нашій планеті. За саме коацерватная крапля, що складається з молекул пептидів, стала основою зародження живого. Це і не викликає сумнівів, адже аналіз внутрішнього складу будь-якого представника біомаси показує, що ці речовини є у всьому: рослинах, тварин, мікроорганізмах, грибах, віруси. Причому вони дуже різноманітні і макромолекулярних по природі.
Назв у цих структур чотири, всі вони є синонімами:
- білки;
- протеїни;
- поліпептиди;
- пептиди.
білкові молекули
Їх кількість воістину незліченно. При цьому всі білкові молекули можна розділити на дві великі групи:
- прості - складаються тільки з амінокислотних послідовностей, сполучених пептидними зв'язками;
- складні - будова і структура білка характеризуються додатковими Протолітична (простетическими) групами, званими ще кофакторами.
При цьому складні молекули також мають свою класифікацію.
Градація складних пептидів
- Глікопротеїди - тісно пов'язані сполуки білка і вуглеводу. У структуру молекули вплітаються простетичноїгрупи мукополісахаридів.
- Ліпопротеїди - комплексне з'єднання з білка і ліпіду.
- Металопротеїни - як простетичної групи виступають іони металів (залізо, марганець, мідь і інші).
- Нуклеопротеїди - зв'язок білка і нуклеїнових кислот (ДНК, РНК).
- Фосфопротеіди - конформація протеїну і залишку ортофосфорної кислоти.
- Хромопротеїди - дуже схожі з Металопротеїни, проте елемент, що входить до складу простетичної групи, являє собою цілий забарвлений комплекс (червоний - гемоглобін, зелений - хлорофіл і так далі).
У кожної розглянутої групи будова і властивості білків різні. Функції, які вони виконують, також варіюються в залежності від типу молекули.

Хімічна будова білків
З цієї точки зору протеїни - це довга, масивна ланцюг амінокислотних залишків, що з'єднуються між собою специфічними зв'язками, званими пептидними. Від бічних структур кислот відходять відгалуження - радикали. Така будова молекули було відкрито Е. Фішером на початку XXI століття.
Пізніше більш детально були вивчені білки, будова і функції білків. Стало ясно, що амінокислот, що утворюють структуру пептиду, всього 20, але вони здатні комбінуватися самим різним способом. Звідси і різноманітність поліпептидних структур. Крім того, в процесі життєдіяльності і виконання своїх функцій білкиздатні зазнавати ряд хімічних перетворень. В результаті вони змінюють структуру, і з'являється вже зовсім новий тип з'єднання.
Щоб розірвати пептидний зв'язок, тобто порушити білок, будова ланцюгів, потрібно підібрати дуже жорсткі умови (дія високих температур, кислот або лугів, каталізатора). Це пояснюється високою міцністю в молекулі, а саме в пептидного групі.

Виявлення білкової структури в умовах лабораторії проводиться за допомогою биуретовой реакції - впливу на поліпептид свіжоосадженого (II). Комплекс пептидного групи і іона міді дає яскраво-фіолетове забарвлення.
Існує чотири основні структурні організації, кожна з яких має свої особливості будови білків.
Рівні організації: первинна структура
Як уже згадувалося вище, пептид - це послідовність амінокислотних залишків з включеннями, коферментами або ж без них. Так ось первинної називають таку структуру молекули, яка є природною, природною, є істинно амінокислоти, з'єднані пептидними зв'язками, і більше нічого. Тобто поліпептид лінійної будови. При цьому особливості будови білків такого плану - в тому, що таке поєднання кислот є визначальним для виконання функцій білкової молекули. Завдяки наявності даних особливостей можливо не тільки ідентифікувати пептид, а й передбачити властивості і роль абсолютно нового, ще не відкритого. Приклади пептидів, що володіють природним первинним будовою, - інсулін, пепсин, хімотрипсин і інші.

вторинна конформація
Будова і властивості білків цієї категорії дещо змінюються. Така структура може сформуватися спочатку від природи або при впливі на первинну жорстким гідролізом, температурою або іншими умовами.
Дана конформація має три різновиди:
- Рівні, правильні, стереорегулярность витки, побудовані із залишків амінокислот, які закручуються навколо центральної осі з'єднання. Утримуються разом лише виникають між киснем однієї пептидного угруповання і воднем інший. Причому будова вважається правильним через те, що витки рівномірно повторюються через кожні 4 ланки. Така структура може бути як левозакрученной, так і правозакрученной. Але в більшості відомих білків переважає правообертальні ізомер. Такі конформації прийнято називати альфа-структурами.
- Склад і будова білків наступного типу відрізняється від попереднього тим, що водневі зв'язки утворюються не між рядом стоять по одній стороні молекули залишками, а між значно віддаленими, причому на досить велику відстань. З цієї причини вся структура набуває вигляду кількох хвилеподібних, звивистих змійкою поліпептидних ланцюжків. Є одна особливість, яку повинен проявляти білок. Будова амінокислот на відгалуженнях повинно бути максимально коротким, як у гліцину або аланіну, наприклад. Цей тип вторинної конформації носить назву бета-листів за здатність ніби злипатися при утворенні загальної структури.
- Що відноситься до третього типу будова білка біологія позначає як складні, разноразбросанние, невпорядковані фрагменти, що не володіють стереорегулярность і здатні змінювати структуру під впливом зовнішніх умов.
Прикладів білків, що мають вторинну структуру від природи, не виявлено.

третинне освіту
Це досить складна конформація, що має назву "кулька". Що собою являє такий білок? Будова його грунтується на вторинній структурі, проте додаються нові типи взаємодій між атомами угруповань, і вся молекула немов згортається, орієнтуючись, таким чином, на те, щоб гідрофільні угруповання були спрямовані всередину глобули, а гідрофобні - назовні.
Цим пояснюється заряд білкової молекули в колоїдних розчинах води. Які ж типи взаємодій тут присутні?
- Водневі зв'язки - залишаються без змін між тими ж самими частинами, що і у вторинній структурі.
- взаємодії - виникають при розчиненні поліпептиду в воді.
- Іонні тяжіння - утворюються між різнозаряджені групами амінокислотних залишків (радикалів).
- Ковалентні взаємодії - здатні формуватися між конкретними кислотними ділянками - молекулами цистеїну, вірніше, їх хвостами.
Таким чином, склад парламенту й будова білків, що володіють третинної структурою, можна описати як згорнуті в глобули поліпептидні ланцюги, які утримують і стабілізуючі свою конформацію за рахунок різних типів хімічних взаємодій. Приклади таких пептидів: фосфогліцераткеназа, тРНК, альфа-кератин, фиброин шовку і інші.
четвертичная структура
Це одна з найскладніших глобул, яку утворюють білки. Будова і функції білків подібного плану дуже багатогранні і специфічні.
Що собою являє така конформація? Це кілька (в деяких випадках десятки) великих і дрібних поліпептидних ланцюгів, які формуються незалежно один від одного. Але потім за рахунок тих же взаємодій, що ми розглядали для третинної структури, всі ці пептиди скручуються і переплітаються між собою. Таким чином виходять складні конформаційні глобули, які можуть містити і атоми металів, і ліпідні угруповання, і вуглеводні. Приклади таких білків: ДНК-полімераза, білкова оболонка тютюнового вірусу, гемоглобін та інші.

Всі розглянуті нами структури пептидів мають свої методи ідентифікації в лабораторних умовах, засновані на сучасних можливостях використання хроматографії, центрифугування, електронної та оптичної мікроскопії і високих комп'ютерних технологіях.
Їх функції
Будова і функції білків тісно корелюють один з одним. Тобто кожен пептид грає певну роль, унікальну і специфічну. Зустрічаються і такі, які здатні виконувати в одній живій клітині відразу кілька значних операцій. Однак можна в узагальненому вигляді висловити основні функції білкових молекул в організмах живих істот:
- Забезпечення руху. Одноклітинні організми, або органели, або деякі види клітин здатні до пересувань, скорочень, переміщенням. Це забезпечується білками, що входять до складу структури їх рухового апарату: війок, джгутиків, цитоплазматичної мембрани. Якщо ж говорити про нездатних до переміщень клітинах, то білки можуть сприяти їх скорочення (міозин м'язів).
- Поживна або резервна функція. Являє собою накопичення білкових молекул в яйцеклітинах, зародках і насінні рослин для подальшого заповнення відсутніх поживних речовин. При розщепленні пептиди дають амінокислоти і біологічно активні речовини, які необхідні для нормального розвитку живих організмів.
- Енергетична функція. Крім вуглеводів, сили організму можуть давати і білки. При розпаді 1 г пептиду вивільняється 17,6 кДж корисної енергії у формі аденозинтрифосфорної кислоти (АТФ), яка витрачається на процеси життєдіяльності.
- Сигнальна і Полягає в здійсненні ретельного контролю за процесами, що відбуваються і передачі сигналів від клітин до тканин, від них до органів, від останніх до систем і так далі. Типовим прикладом може служити інсулін, який строго фіксує кількість глюкози в крові.
- Рецепторная функція. Здійснюється шляхом зміни конформації пептиду з одного боку мембрани і залучення в реструктуризацію іншого кінця. При цьому і відбувається передача сигналу і необхідної інформації. Найчастіше такі білки вбудовуються в цитоплазматические мембрани клітин і здійснюють суворий контроль над усіма речовинами, що проходять через неї. Також сповіщають про хімічних і фізичних змінах навколишнього середовища.
- Транспортна функція пептидів. Її здійснюють білки-канали і білки-переносники. Роль їх очевидна - транспортування необхідних молекул до місць з низькою концентрацією з частин з високою. Типовим прикладом служить перенесення кисню і діоксиду вуглецю по органам і тканинам білком гемоглобіном. Ними ж здійснюється доставка з'єднань з невисокою молекулярною масою через мембрану клітини всередину.
- Структурна функція. Одна з найважливіших з тих, які виконує білок. Будова всіх клітин, їх органел забезпечується саме пептидами. Вони подібно каркасу задають форму і структуру. Крім того, вони ж її підтримують і видозмінюють в разі потреби. Тому для зростання і розвитку всіх живих організмів необхідні білки в раціоні харчування. До таких пептидів можна віднести еластин, тубулін, колаген, актин, кератин і інші.
- Каталітична функція. Її виконують ферменти. Численні і різноманітні, вони прискорюють всі хімічні і біохімічні реакції в організмі. Без їх участі звичайне яблуко в шлунку змогло б перетравитися тільки за два дні, з великою ймовірністю загнити при цьому. Під дією каталази, пероксидази та інших ферментів цей процес відбувається за дві години. В цілому саме завдяки такій ролі білків здійснюється анаболізм і катаболізм, тобто пластичний і

Захисна роль
Існує кілька типів загроз, від яких білки покликані оберігати організм.
По-перше, травмуючих реагентів, газів, молекул, речовин різного спектру дії. Пептиди здатні вступати з ними в хімічну взаємодію, переводячи в нешкідливу форму або ж просто нейтралізуючи.
По-друге, фізична загроза з боку ран - якщо білок фібриноген вчасно не трансформується в фібрин на місці травми, то кров не згорнеться, а значить, закупорка не відбудеться. Потім, навпаки, знадобиться пептид плазмин, здатний згусток розсмоктати і відновити прохідність судини.
По-третє, загроза імунітету. Будова і значення білків, що формують імунний захист, вкрай важливі. Антитіла, імуноглобуліни, інтерферони - все це важливі і значущі елементи лімфатичної та імунної системи людини. Будь-яка чужорідна частинка, шкідлива молекула, відмерла частина клітини або ціла структура піддається негайному дослідженню з боку пептидного сполучення. Саме тому людина може самостійно, без допомоги лікарських засобів, щодня захищати себе від інфекцій і нескладних вірусів.
Фізичні властивості
Будова білка клітини досить специфічний і залежить від виконуваної функції. А ось фізичні властивості всіх пептидів схожі і зводяться до наступних характеристиках.
- Вага молекули - до 1000000 Дальтон.
- У водному розчині формують колоїдні системи. Там структура набуває заряд, здатний варіюватися в залежності від кислотності середовища.
- При впливі жорстких умов (опромінення, кислота або луг, температура і так далі) здатні переходити на інші рівні конформаций, тобто денатурувати. Даний процес в 90% випадків є незворотнім. Однак існує і зворотний зсув - ренатурації.
Це основні властивості фізичної характеристики пептидів.
В основі життєдіяльності клітини лежать біохімічні процеси, що протікають на молекулярному рівні і службовці предметом вивчення біохімії. Відповідно і явища спадковості і мінливості теж пов'язані з молекулами органічних речовин, і в першу чергу з нуклеїновими кислотами і білками.
склад білків
Білки являють собою великі молекули, що складаються з сотень і тисяч елементарних ланок - амінокислот. Такі речовини, що складаються з повторюваних елементарних ланок - мономерів, називаються полімерами. Відповідно білки можна назвати полімерами, мономерами яких служать амінокислоти.
Всього в живій клітині відомо 20 видів амінокислот. Назва амінокислоти отримали з-за змісту в своєму складі аминной групи NHy, що володіє основними властивостями, і карбоксильної групи СООН, що має кислотні властивості. Всі амінокислоти мають однакову групу NH2-СН-СООН і відрізняються один від одного хімічною групою, званої радикалом - R. З'єднання амінокислот в полімерну ланцюг відбувається завдяки освіті пептидного зв'язку (СО - NH) між карбоксильною групою однієї амінокислоти і аміногрупою іншої амінокислоти. При цьому виділяється молекула води. Якщо утворилася полімерна ланцюг коротка, вона називається олігопептідной, якщо довга - поліпептидного.
будова білків
При розгляді будови білків виділяють первинну, вторинну, третинну структури.
первинна структура визначається порядком чергування амінокислот в ланцюзі. Зміна в розташуванні навіть однієї амінокислоти веде до утворення абсолютно нової молекули білка. Число білкових молекул, яке утворюється при поєднанні 20 різних амінокислот, досягає астрономічної цифри.
Якби великі молекули (макромолекули) білка розташовувалися в клітці в витягнутому стані, вони займали б у ній занадто багато місця, що ускладнило б життєдіяльність клітини. У зв'язку з цим молекули білка скручуються, згинаються, згортаються в самі різні конфігурації. Так на основі первинної структури виникає вторинна структура - білкова ланцюг укладається в спіраль, що складається з рівномірних витків. Сусідні витки з'єднані між собою слабкими водневими зв'язками, які при багаторазовому повторенні надають стійкість молекулам білків з цією структурою.
Спіраль вторинної структури укладається в клубок, утворюючи третинну структуру. Форма клубка у кожного виду білків строго специфічна і повністю залежить від первинної структури, т. Е. Від порядку розташування амінокислот в ланцюзі. Третинна структура утримується завдяки безлічі слабких електростатичних зв'язків: позитивно і негативно заряджені групи амінокислот притягуються і зближують навіть далеко віддалені один від одного ділянки білкової ланцюга. Зближуються і інші ділянки білкової молекули, що несуть, наприклад, гідрофобні (водовідштовхувальні) групи.
Деякі білки, наприклад гемоглобін, складаються з декількох ланцюгів, що розрізняються по первинній структурі. Об'єднуючись разом, вони створюють складний білок, що володіє не тільки третинної, а й четвертичной структурою (Рис. 2).
У структурах білкових молекул спостерігається наступна закономірність: чим вище структурний рівень, тим слабкіше підтримують їх хімічні зв'язки. Зв'язки, що утворюють четвертичную, третинну, вторинну структуру, вкрай чутливі до фізико-хімічних умов середовища, температури, радіації і т. Д. Під їх впливом структури молекул білків руйнуються до первинної - вихідної структури. Таке порушення природної структури білкових молекул називається денатурацією.При видаленні денатурирующего агента багато білків здатні мимовільно відновлювати вихідну структуру. Якщо ж природний білок піддається дії вьюокой температури або інтенсивного дії інших факторів, то він необоротно денатурируется. Саме фактом наявності незворотною денатурації білків клітин пояснюється неможливість життя в умовах дуже високої температури.



Біологічна роль білків в клітині
Білки, звані також протеїнами (Грец. Протос - перший), в клітинах тварин і рослин виконують різноманітні і дуже важливі функції, до яких можна віднести наступні.
Каталітична. Природні каталізатори - ферменти являють собою повністю або майже повністю білки. Завдяки ферментам хімічні процеси в живих тканинах прискорюються в сотні тисяч або в мільйони разів. Під їх дією всі процеси йдуть миттєво в «м'яких» умовах: при нормальній температурі тіла, в нейтральній для живої тканини середовищі. Швидкодія, точність і вибірковість ферментів непорівнянні ні з одним з штучних каталізаторів. Наприклад, одна молекула ферменту за одну хвилину здійснює реакцію розпаду 5 млн. Молекул пероксиду водню (Н202). Ферментам характерна вибірковість. Так, жири розщеплюються спеціальним ферментом, який не діє на білки і полісахариди (крохмаль, глікоген). У свою чергу, фермент, який розщеплює тільки крохмаль або глікоген, не діє на жири.
Процес розщеплення чи синтезу будь-якого речовини в клітині, як правило, розділений на ряд хімічних операцій. Кожну операцію виконує окремий фермент. Група таких ферментів становить біохімічний конвейер.
Вважають, що каталітична функція білків залежить від їх третинної структури, при її руйнуванні каталітична активність ферменту зникає.
Захисна. Деякі види білків захищають клітину і в цілому організм від попадання в них хвороботворних мікроорганізмів і чужорідних тіл. Такі білки звуться антитіл.Антитіла зв'язуються з чужорідними для організму білками бактерій і вірусів, що пригнічує їх розмноження. На кожен чужорідний білок організм виробляє спеціальні «антібелкі» - антитіла. Такий механізм опору збудників захворювань називається імунітетом.
Щоб попередити захворювання, людям і тваринам вводять ослаблені або вбиті збудники (вакцини), які не викликають хворобу, але змушують спеціальні клітини організму виробляти антитіла проти цих збудників. Якщо через деякий час хвороботворні віруси і бактерії потрапляють в такий організм, вони зустрічають міцний захисний бар'єр з антитіл.
Гормональна. Багато гормони також є білки. Поряд з нервовою системою гормони керують роботою різних органів (і всього організму) через систему хімічних реакцій.
Відбивна. Білки клітини здійснюють прийом сигналів, що йдуть ззовні. При цьому різні фактори середовища (температурний, хімічний, механічний і ін.) Викликають зміни в структурі білків - оборотну денатурацію, яка, в свою чергу, сприяє виникненню хімічних реакцій, що забезпечують відповідь клітини на зовнішнє роздратування. Ця здатність білків лежить в основі роботи нервової системи, мозку.
Рухова. Всі види рухів клітини і організму: мерехтіння війок у найпростіших, скорочення м'язів у вищих тварин і інші рухові процеси - виробляються особливим видом білків.
Енергетична. Білки можуть служити джерелом енергії для клітини. При нестачі вуглеводів або жирів окислюються молекули амінокислот. Звільнена у своїй енергія використовується для підтримки процесів життєдіяльності організму.
Транспортна. Білок гемоглобін крові здатний зв'язувати кисень повітря і транспортувати його по всьому тілу. Ця найважливіша функція властива і деяким іншим білкам.
Пластична. Білки - основний будівельний матеріал клітин (їх мембран) і організмів (їх кровоносних судин, нервів, травного тракту і ін.). При цьому білки володіють індивідуальною специфічністю, т. Е. В організмах окремих людей містяться деякі, характерні лише для нього, білки
Таким чином, білки - ці найважливіший компонент клітини, без якого неможливо прояв властивостей життя. Однак відтворення живого, явищеспадковості, як ми побачимо пізніше, пов'язане з молекулярними структурами нуклеїнових кислот. Це відкриття - результат новітніх досягнень біології. Тепер відомо, що жива клітина обов'язково володіє двома видами полімерів-білками і нуклеїновими кислотами. В їх взаємодії укладені найглибші боку явища життя.
1. Чому білки вважаються полімерами?
Відповідь. Білки - це полімери, тобто молекули, побудовані, як ланцюга, з повторюваних мономірних ланок, або субодиниць, що складаються з амінокислот, з'єднаних в певній послідовності пептидного зв'язком. Вони - основні і необхідні складові частини всіх організмів.
Розрізняють білки прості (протеїни) і складні (протеїди). Протеїни - білки, молекули яких містять тільки білкові компоненти. При повному їх гідролізі утворюються амінокислоти.
Протеідамі називають складні білки, молекули яких істотно відрізняються від молекул протеїнів тим, що крім власне білкового компонента містять низькомолекулярний компонент небілкової природи
2. Які функції білків вам відомі?
Відповідь. Білки виконують такі функції: будівельну, енергетичну, каталітичну, захисну, транспортну, скоротливу, сигнальну і інші.
Питання після § 11
1. Які речовини називаються білками?
Відповідь. Білки, або протеїни, являють собою біологічні полімери, мономерами яких є амінокислоти. Всі амінокислоти мають аміногрупу (-NH2) і карбоксильну групу (-СООН) і розрізняються будовою і властивостями радикалів. Амінокислоти пов'язані між собою пептидними зв'язками, тому білки називають ще полипептидами.
Відповідь. Молекули білків можуть приймати різні просторові форми - конформації, які представляють собою чотири рівні їх організації. Лінійна послідовність амінокислот у складі поліпептидного ланцюга являє первинну структуру білка. Вона унікальна для будь-якого білка і визначає його форму, властивості і функції.
3. Як утворюються вторинна, третинна і четвертинна структури білка?
Відповідь. Вторинна структура білка формується при утворенні водневих зв'язків між -СО- і -NH- групами. При цьому поліпептидний ланцюг закручується в спіраль. Спіраль може набувати конфігурацію глобули, так як між радикалами амінокислот в спіралі виникають різноманітні зв'язки. Глобула - третинна структура білка. Якщо кілька глобул об'єднуються в єдиний комплекс, то виникає четвертичная структура. Наприклад, гемоглобін крові людини утворений чотирма глобулами.
4. Що таке денатурація білка?
Відповідь. Порушення природної структури білка називається денатурацією. Під дією ряду факторів (хімічних, радіоактивних, температурних та ін.) Може зруйнуватися четвертичная, третинна і вторинна структури білка. У разі, якщо дія фактора припиняється, білок може відновити свою структуру. Якщо ж дія фактора наростає, руйнується і первинна структура білка - поліпептидний ланцюг. Це вже незворотній процес - відновити структуру білок не може
5. За якою ознакою білки діляться на прості і складні?
Відповідь. Прості білки складаються виключно з амінокислот. До складу складних білків можуть входити інші органічні речовини: вуглеводи (тоді вони називаються гликопротеинами), жири (ліпопротеїни), нуклеїнові кислоти (нуклеопротеїнами).
6. Які функції білків вам відомі?
Відповідь. Будівельна (пластична) функція. Білки є структурним компонентом біологічних мембран і органоїдів клітини, а також входять до складу опорних структур організму, волосся, нігтів, судин. Ферментативна функція. Білки є ферментами, т. Е. Біологічними каталізаторами, які прискорюють швидкість біохімічних реакцій в десятки і сотні мільйонів разів. Прикладом може служити амілаза, що розщеплює крохмаль до моносахаридів. Скорочувальна (рухова) функція. Її виконують особливі скоротливі білки, що забезпечують рух клітин і внутрішньоклітинних структур. Завдяки їм переміщуються хромосоми при діленні клітини, а джгутики і війки надають руху клітини найпростіших. Скоротливі властивості білків актину і міозину лежать в основі роботи м'язів. Транспортна функція. Білки беруть участь в транспорті молекул і іонів в межах організму (гемоглобін переносить кисень з легень до органів і тканин, альбумін сироватки крові бере участь в транспорті жирних кислот). Захисна функція. Вона полягає в запобіганні організму від пошкоджень і вторгнення чужорідних білків і бактерій. Білки-антитіла, що виробляються лімфоцитами, створюють захист організму від чужорідної інфекції, тромбін і фібрин беруть участь в утворенні тромбу, тим самим допомагаючи організму уникнути великих втрат крові. Регуляторна функція. Її виконують білки-гормони. Вони беруть участь в регуляції активності клітини і всіх життєвих процесів організму. Так, інсулін регулює рівень цукру в крові і підтримує його на певному рівні. Сигнальна функція. Білки, вбудовані в мембрану клітини, здатні змінювати свою структуру у відповідь на роздратування. Тим самим передаються сигнали із зовнішнього середовища всередину клітини. Енергетична функція. Вона реалізується білками вкрай рідко. При повному розщеплюванні 1 г білка здатне виділитися 17,6 кДж енергії. Однак білки для організму - дуже цінне з'єднання. Тому розщеплення білка відбувається зазвичай до амінокислот, з яких будуються нові поліпептидні ланцюжки. Білки-гормони регулюють активність клітини і всіх життєвих процесів організму. Так, в організмі людини соматотропин бере участь в регуляції росту тіла, інсулін підтримує на постійному рівні вміст глюкози в крові.
7. Яку роль виконують білки-гормони?
Відповідь. Регуляторна функція властива білкам-гормонів (регуляторам). Вони регулюють різні фізіологічні процеси. Наприклад, найбільш відомим гормоном є інсулін, який регулює вміст глюкози в крові. При нестачі інсуліну в організмі виникає захворювання, відоме як цукровий діабет.
8. Яку функцію виконують білки-ферменти?
Відповідь. Ферменти є біологічними каталізаторами, тобто прискорювачами хімічних реакцій в сотні мільйонів разів. Ферменти мають сувору специфічністю по відношенню до речовини, що вступає в реакцію. Кожна реакція каталізується своїм ферментом.
9. Чому білки рідко використовуються в якості джерела енергії?
Відповідь. Мономери білків амінокислоти - цінна сировина для побудови нових білкових молекул. Тому повне розщеплення поліпептидів до неорганічних речовин відбувається рідко. Отже, енергетична функція, яка полягає у виділенні енергії при повному розщепленні, виконується білками досить рідко.
Білок яйця є типовим протеїном. З'ясуйте, що з ним станеться, якщо на нього подіяти водою, спиртом, ацетоном, кислотою, лугом, рослинним маслом, високою температурою і т. Д.
Відповідь. В результаті дії високої температури на білок яйця відбудеться денатурація білка. При дії спирту, ацетону, кислотами або лугами відбувається приблизно те ж саме: білок згортається. Це процес, при якому відбувається порушення третинної і четвертинної структури білка внаслідок розриву водневих і іонних зв'язків.
У воді і рослинному маслі білок зберігає свою структуру.
Подрібніть бульба сирої картоплі до стану кашки. Візьміть три пробірки і в кожну покладіть невелику кількість подрібненого картоплі.
Першу пробірку помістіть в морозилку холодильника, другу - на нижню полицю холодильника, а третю - в банку з теплою водою (t \u003d 40 ° С). Через 30 хв дістаньте пробірки і в кожну капніть невелику кількість пероксиду водню. Поспостерігайте, що буде відбуватися в кожній пробірці. Поясніть отримані результати
Відповідь. Даний досвід ілюструє активність ферменту каталази живої клітини на перекис водню. В результаті реакції виділяється кисень. За динамікою виділення бульбашок можна судити про активність ферменту.
Досвід дозволив зафіксувати наступні результати:
Активність каталази залежить від температури:
1. Пробірка 1: бульбашок немає - це тому, при низькій температурі клітини картоплі зруйнувалися.
2. Колба 2: бульбашок невелика кількість - тому, що активність ферменту при низькій температурі невисока.
3. Пробірка 3: бульбашок багато, температура оптимальна, каталаза дуже активна.
В першу пробірку з картоплею капніть кілька крапель води, в другу - кілька крапель кислоти (столовий оцет), а в третю - лугу.
Поспостерігайте, що буде відбуватися в кожній пробірці. Поясніть отримані результати. Зробіть висновки.
Відповідь. При додаванні води нічого не відбувається, при додаванні кислоти відбувається деяке потемніння, при додаванні лугу «вспенивание» - лужний гідроліз.
Схожі статті
-
В якій послідовності краще читати Террі Пратчетта, з чого почати?
«Це б-ла нас-то-ящая га-Зета з 96 стра-ка-ми част-них об'єк-яв-ле-ний. Імен-но там я ви-вчив-ся азам мас-терс-тва, трю-кам, саль-ним шу-точ-кам, сом-ні-тель-но-му біт-кло-ру і штам-пам ре- ги-Онал-ної жур-на-лис-ти-ки. І це б-ло ...
-
Планета Меркурій - найближча до Сонця Атмосфера меркурій на 99 складається з
Зараз набула широкого розповсюдження-страненіе ідея про те, що Меркурій був колись супутником Венери. Ця гіпотеза народилася в кінці XIX в. До гіпотезі не сто-сілісь серйозно до тих пір, поки перші польоти космічних апаратів до Меркурія ні ...
-
Кіберкуб - перший крок в четвертий вимір 4 х мірний куб gif
В геометрії гиперкуб - це n-мірному аналогія квадрата (n \u003d 2) і куба (n \u003d 3). Це замкнута опукла фігура, що складається з груп паралельних ліній, розташованих на протилежних краях фігури, і з'єднаних один з одним під прямим ...
-
Хто такі масони? Масони: хто вони? Що значить Массон
Уже в новий час масони створили тексти, де доводили давнє походження свого ордена. Якщо поцікавитися, хто такі масони і чим вони займаються, можна помітити, що вони серйозно відрізняються від своїх попередників. Перші ...
-
Найважливіші правила поведінки для учнів в школі Правило поведінки як вести в школі
Сучасний етикет - це цілий комплекс правил поведінки і хороших манер, який вчить, як треба знайомитися, вітатися, як вести себе в громадських місцях, як ходити в гості, як правильно накривати на стіл і вести себе під час ...
-
Українська історія і російська весна Російська весна вікі
Є серйозні підозри, що сайт «Російська весна» rusvesna.su є американським провокаційним проектом.Сам сайт якісний. Добірка новин теж хороша, але є серйозні насторожуючі моменти.1. Абсолютно незрозуміло хто стоїть за ...