Молекула ДНК. Структура молекули ДНК. Спадкова інформація: зберігання і передача. Генетичний код. Ланцюжок ДНК Де містяться молекули ДНК
Нуклеїнові кислоти - високомолекулярні речовини, що складаються з мононуклеотидів, які з'єднані один з одним в полімерну ланцюжок за допомогою 3 ", 5" - фосфодіефірних зв'язків і упаковані в клітинах певним чином.
Нуклеїнові кислоти - біополімери двох різновидів: рибонуклеїнова кислота (РНК) і дезоксирибонуклеїнової кислоти (ДНК). Кожен біополімер складається з нуклеотидів, що розрізняються по вуглеводному залишку (рибозе, дезоксирибози) і одному з азотистих основ (урацил, тимін). Відповідно до цих відмінностей нуклеїнові кислоти і отримали свою назву.
Структура дезоксирибонуклеїнової кислоти
Нуклеїнові кислоти мають первинну, вторинну і третинну структуру.
Первинна структура ДНК
Первинною структурою ДНК називають лінійну полінуклеотидних ланцюг, в якій мононуклеотиди з'єднані 3 ", 5" -фосфодіефірнимі зв'язками. Вихідним матеріалом при складанні ланцюга нуклеїнової кислотив клітці є нуклеозид 5 "-тріфосфат, який в результаті видалення β і γ залишків фосфорної кислоти здатний приєднати 3" -атом вуглецю іншого нуклеозиду. Таким чином, 3 "-атом вуглецю однієї дезоксирибози ковалентно зв'язується з 5" -атомом вуглецю інший дезоксирибози за допомогою одного залишку фосфорної кислоти і утворює лінійну полінуклеотидних ланцюг нуклеїнової кислоти. Звідси і назва: 3 ", 5" -фосфодіефірние зв'язку. Азотисті основи не беруть участі в з'єднанні нуклеотидів одного ланцюга (рис. 1.).
Таке з'єднання, між залишком молекули фосфорної кислоти одного нуклеотиду і вуглеводом іншого, призводить до утворення пентозо-фосфатного кістяка молекули полинуклеотида, на якому збоку один за іншим приєднуються азотисті основи. Їх послідовність розташування в ланцюгах молекул нуклеїнових кислот строго специфічна для клітин різних організмів, тобто носить видовий характер (правило Чаргаффа).
Лінійна ланцюг ДНК, довжина якої залежить від числа входять до ланцюг нуклеотидів, має два кінці: один називається 3 "-кінців і містить вільний гідроксил, а інший - 5"-кінців, містить залишок фосфорної кислоти. Ланцюг полярна і може мати напрвление 5 "-> 3" і 3 "-> 5". Винятком є кільцеві ДНК.
Генетичний "текст" ДНК складений за допомогою кодових "слів" - триплетів нуклеотидів, званих кодонами. Ділянки ДНК, що містять інформацію про первинну структуру всіх типів РНК, називають структурними генами.
Полінуклеодітние ланцюжка ДНК досягають гігантських розмірів, тому в клітці вони упаковані певним чином.
Вивчаючи склад ДНК, Чаргафф (1949) встановив важливі закономірності, що стосуються змісту окремих підстав ДНК. Вони допомогли розкрити вторинну структуру ДНК. Ці закономірності називають правилами Чаргаффа. Правила Чаргаффа
Ці правила говорять про те, що при побудові ДНК повинна дотримуватися досить суворе відповідність (спаровування) НЕ пуринових і піримідинових основ взагалі, а конкретно тиміну з аденін і цитозину з гуаніном. На підставі цих правил в тому числі, в 1953 р Уотсон і Крик запропонували модель вторинної структури ДНК, що отримала назву подвійної спіралі (рис.). |

Вторинна структура ДНК
Вторинна структура ДНК - це подвійна спіраль, модель якої була запропонована Д. Уотсон і Ф. Криком в 1953 році.
Передумови до створення моделі ДНК
В результаті первинних аналізів склалося уявлення, що ДНК будь-якого походження містить всі чотири нуклеотиду в рівних молярних кількостях. Однак в 1940-х роках Е. Чаргафф і його співробітники в результаті аналізу ДНК, виділених з різноманітних організмів, ясно показали, що азотисті основи містяться в них в різних кількісних співвідношеннях. Чаргафф знайшов, що, хоча ці співвідношення однакові для ДНК з усіх клітин одного і того ж виду організмів, ДНК від різних видів можуть помітно відрізнятися за змістом тих чи інших нуклеотидів. Це наводило на думку, що відмінності в співвідношенні азотистих основ, можливо, пов'язані з якимось біологічним кодом. Хоча співвідношення окремих пуринових і піримідинових основ у різних зразках ДНК виявилося неоднаковим, при порівнянні результатів аналізів виявилася певна закономірність: у всіх зразках загальна кількість пуринів було дорівнює загальній кількості пиримидинов (А + Г = Т + Ц), кількість аденіну - кількості тиміну (А = Т), а кількість гуаніну - кількості цитозину (Г = Ц). ДНК, виділена з клітин ссавців, була в цілому багатшими аденином і тиміном і щодо біднішими гуаніном і цитозином, тоді як у бактерій ДНК була багатшою гуаніном і цитозином і щодо біднішими аденином і тиміном. Ці дані склали важливу частину фактичного матеріалу, на основі якого пізніше була побудована модель структури ДНК Уотсона - Кріка.
Ще одним важливим непрямим свідченням можливу структуру ДНК послужили дані Л. Полінга про будову білкових молекул. Полінг показав, що можливо кілька різних стійких конфігурацій амінокислотної ланцюга в білкової молекулі. Одна з поширених конфігурацій пептидного ланцюга - α-спіраль - являє собою правильну гвинтоподібну структуру. При такій структурі можливе утворення водневих зв'язків між амінокислотами, які перебувають на суміжних витках ланцюга. Полінг описав α-спіральну конфігурацію поліпептидного ланцюга в 1950 році і висловив припущення, що і молекули ДНК, ймовірно, мають спіральну структуру, закріплену водневими зв'язками.
Однак найбільш цінні відомості про будову молекули ДНК дали результати рентгеноструктурного аналізу. Рентгенівські промені, проходячи крізь кристал ДНК, зазнають дифракцію, т. Е. Відхиляються в певних напрямках. Ступінь і характер відхилення променів залежать від структури самих молекул. Дифракційна рентгенограма (рис. 3) дає досвідченому оку ряд непрямих вказівок щодо будови молекул досліджуваної речовини. Аналіз дифракційних рентгенограм ДНК привів до висновку, що азотисті основи (що лежать рівно) укладені на зразок стопки тарілок. Рентгенограми дозволили виявити в структурі кристалічної ДНК три головні періоди: 0,34, 2 і 3,4 нм.

Модель ДНК Уотсона-Кріка
Виходячи з аналітичних даних Чаргаффа, рентгенограм, отриманих Вілкінсом і досліджень хіміків, які надали відомості про точні відстанях між атомами в молекулі, про кути між зв'язками даного атома і про величину атомів, Уотсон і Крик почали будувати фізичні моделі окремих складових частин молекули ДНК в певному масштабі і "підганяти" їх один до одного з таким розрахунком, щоб отримана система відповідала різним експериментальним даними [Показати] .
Ще раніше було відомо, що в ланцюзі ДНК сусідні нуклеотиди з'єднані фосфодіефірнимі містками, що зв'язують 5 "-вуглецевого атом дезоксирибози одного нуклеотиду з 3" -вуглецевим атомом дезоксирибози наступного нуклеотиду. Уотсон і Крик не сумнівалися в тому, що період 0,34 нм відповідає відстані між послідовними нуклеотидами в ланцюжку ДНК. Далі, можна було припускати, що період 2 нм відповідає товщині ланцюга. А для того щоб пояснити, яку реальну структурі відповідає період 3,4 нм, Уотсон і Крик, так само як раніше Полінг, припустили, що ланцюг закручена у вигляді спіралі (або, точніше, утворює кручені лінію, так як спіраль в строгому сенсі цього слова виходить тоді, коли витки утворюють в просторі конічну, а не циліндричну поверхню). Тоді період 3,4 нм буде відповідати відстані між послідовними витками цієї спіралі. Така спіраль може бути дуже щільною або ж кілька розтягнутої, т. Е. Витки її можуть бути пологими або крутими. Оскільки період 3,4 нм рівно в 10 разів більше відстані між послідовними нуклеотидами (0,34 нм), ясно, що кожен повний виток спіралі містить 10 нуклеотидів. За цими даними Уотсон і Крик змогли обчислити щільність полінуклеотидних ланцюга, закрученої в спіраль діаметром 2 нм, з відстанню між витками, рівним 3,4 нм. Виявилося, що у такому колі щільність була б удвічі менше фактичної щільності ДНК, яка була вже відома. Довелося припустити, що молекула ДНК складається з двох ланцюгів - що це подвійна спіраль з нуклеотидів.
Наступним завданням було, звичайно, з'ясування просторових відносин між обома ланцюгами, що утворюють подвійну спіраль. Випробувавши на своїй фізичній моделі ряд варіантів розташування ланцюгів, Уотсон і Крик знайшли, що всім наявним даними найкраще відповідає такий варіант, в якому дві полінуклеотидні спіралі йдуть в протилежних напрямках; при цьому ланцюзі, що складаються із залишків цукру і фосфату, утворюють поверхню подвійної спіралі, а пуринів і піримідинів розташовуються усередині. Розташовані один проти одного підстави, що належать двом ланцюгах, попарно з'єднані водневими зв'язками; саме ці водневі зв'язку і утримують ланцюга разом, фіксуючи таким чином загальну конфігурацію молекули.
Подвійну спіраль ДНК можна уявити собі у вигляді спіралі закрученої мотузкових сходів, так щоб поперечини її залишалися в горизонтальному положенні. Тоді дві поздовжні мотузки будуть відповідати ланцюгах із залишків цукру і фосфату, а поперечини - парам азотистих основ, з'єднаних водневими зв'язками.
В результаті подальшого вивчення можливих моделей Уотсон і Крик прийшли до висновку, що кожна "перекладина" повинна складатися з одного пурину і одного пиримидина; при періоді 2 нм (що відповідає діаметру подвійної спіралі) для двох пуринів не вистачило б місця, а два пиримидина не могли б при цьому розташовуватися досить близько один до одного, щоб утворити відповідні водневі зв'язку. Поглиблене дослідження детальної моделі показало, що аденін і цитозин, складаючи відповідну за розмірами комбінацію, все ж не могли б розташовуватися таким чином, щоб між ними утворилися водневі зв'язку. Аналогічні повідомлення змусили виключити також комбінацію гуанін - тимін, тоді як поєднання аденін - тимін і гуанін - цитозин виявилися цілком прийнятними. Природа водневих зв'язків така, що аденін утворює пару з тиміном, а гуанін - з цитозином. Це уявлення про специфічний спарюванні підстав дозволяло пояснити "правило Чаргаффа", згідно з яким в будь-який молекулі ДНК кількість аденіну завжди дорівнює змісту тиміну, а кількість гуаніну - кількості цитозину. Між аденином і тиміном утворюються дві водневі зв'язку, а між гуаніном і цитозином - три. Завдяки цій специфічності в утворенні водневих зв'язків проти кожного аденіну в одного ланцюга в інший виявляється тимін; точно так же проти кожного гуаніну може перебувати тільки цитозин. Таким чином, ланцюги комплементарні один одному, т. Е. Послідовність нуклеотидів в одного ланцюга однозначно визначає їх послідовність в іншій. Дві ланцюга йдуть в протилежних напрямках, і їх кінцеві фосфатні групи знаходяться на протилежних кінцях подвійної спіралі.
В результаті своїх досліджень, в 1953 році Уотсон і Крик запропонували модель будови молекули ДНК (рис. 3), яка залишається актуальною по теперішній час. Згідно з моделлю молекула ДНК складається з двох комплементарних полінуклеотидних ланцюгів. Кожна ланцюг ДНК являє полинуклеотид, що складається з декількох десятків тисяч нуклеотидів. У ній сусідні нуклеотиди утворюють регулярний пентозо-фосфатний кістяк за рахунок з'єднання залишку фосфорної кислоти і дезоксирибози міцної ковалентного зв'язком. Азотисті основи одного полінуклеотидних ланцюга при цьому розташовуються в строго визначеному порядку проти азотистих основ інший. Чергування азотистих основ в полінуклеотидних ланцюга нерегулярно.
Розташування азотистих основ в ланцюзі ДНК є комплементарним (від грец. "Комплемент" - доповнення), тобто проти аденіну (А) завжди виявляється тимін (Т), а проти гуаніну (Г) - тільки цитозин (Ц). Це пояснюється тим, що А і Т, а також Г і Ц строго відповідають один одному, тобто доповнюють один одного. Таке відповідність задається хімічною структуроюпідстав, що дозволяє утворити водневі зв'язки в парі пурину і піримідину. Між А і Т виникають дві зв'язку, між Г і Ц - три. Ці зв'язки забезпечують часткову стабілізацію молекули ДНК в просторі. Стійкість подвійної спіралі при цьому прямо пропорційна числу зв'язків G≡С, є більш стабільними в порівнянні зі зв'язками А = Т.
Відома послідовність розташування нуклеотидів в одного ланцюга ДНК дозволяє за принципом комплементарності встановити нуклеотиди іншого ланцюга.
Крім того, встановлено, що азотисті основи, які мають ароматичну структуру, в водному розчинірозташовуються один над іншим, формуючи як би стопку монет. Такий процес формування стопок з органічних молекул називається стекінг. Полінуклеотидні ланцюга молекули ДНК розглянутій моделі Уотсона-Кріка мають аналогічне фізико-хімічний стан, їх азотисті основи розташовуються у вигляді стопки монет, між площинами яких виникають ван-дер-ваальсові взаємодії (стекінг-взаємодії).
водневі зв'язкуміж комплементарними підставами (по горизонталі) і стекінг-взаємодія між площинами підстав в полінуклеотидних ланцюга за рахунок ван дер Ваальсових сил (по вертикалі) забезпечує молекулі ДНК додаткову стабілізацію в просторі.
Сахарофосфатнимі остови обох ланцюгів звернені назовні, а підстави всередину, назустріч один одному. Напрямок ланцюгів в ДНК антипараллельно (одна з них має напрямок 5 "-> 3", інша - 3 "-> 5", тобто 3 "-кінець одного ланцюга розташований навпроти 5"-кінців інший.). Ланцюги утворюють праві спіралі із загальною віссю. Один виток спіралі становить 10 нуклеотидів, розмір витка 3,4 нм, висота кожного нуклеотиду 0,34 нм, діаметр спіралі - 2,0 нм. В результаті обертання одного ланцюга навколо іншої, утворюється велика борозна (діаметром близько 20 Å) і мала борозна (близько 12 Å) подвійної спіралі ДНК. Така форма подвійної спіралі Уотсона-Кріка в подальшому отримала назву В-форми. У клітинах ДНК зазвичай існує в В-формі, яка є найстабільнішою.
функції ДНК
Запропонована модель пояснювала багато біологічні властивостідезоксирибонуклеїнової кислоти, в тому числі зберігання генетичної інформації і різноманіття генів, що забезпечується великою різноманітністю послідовних поєднань 4-х нуклеотидів і фактом існування генетичного коду, здатність до самовідтворення і передачі генетичної інформації, що забезпечується процесом реплікації, і реалізацію генетичної інформації у вигляді білків, а також будь-яких інших сполук, що утворюються за допомогою білків-ферментів.
Oсновной функції ДНК.
- ДНК є носієм генетичної інформації, що забезпечується фактом існування генетичного коду.
- Відтворення і передана генетичної інформації в поколіннях клітин і організмів. Ця функція забезпечується процесом реплікації.
- Реалізація генетичної інформації у вигляді білків, а також будь-яких інших сполук, що утворюються за допомогою білків-ферментів. Ця функція забезпечується процесами транскрипції і трансляції.

Форми організації двухцепочечной ДНК
ДНК може формувати кілька типів подвійних спіралей (рис.4). В даний час вже відомо шість форм (від А до Е і Z-форма).
Структурні форми ДНК, як встановила Розалінд Франклін, залежать від насичення водою молекули нуклеїнової кислоти. У дослідженнях волокон ДНК за допомогою рентгеноструктурного аналізу було показано, що рентгенограма радикальним чином залежить від того, при якій відносної вологості, при якій мірі насичення водою цього волокна відбувається експеримент. Якщо волокно було досить насичене водою, то виходила одна рентгенограма. При висушуванні виникала зовсім інша рентгенограма, сильно відрізняється від рентгенограми волокна високої вологості.
Молекула ДНК високої вологості отримала назву В-форми. При фізіологічних умовах (низька концентрація солі, висока степере гідратації) домінуючим структурним типом ДНК є По-форма (основна форма двухцепочечной ДНК - модель Уотсона-Кріка). Крок спіралі такий молекули дорівнює 3,4 нм. На виток припадає 10 комплементарних пар у вигляді скручених стопок "монет" - азотистих основ. Стопки утримуються водневими зв'язками між двома протилежними "монетами" стопок, і "обмотані" двома стрічками фосфодіефірнимі остова, закрученими в праву спіраль. Площині азотистих основ перпендикулярні осі спіралі. Сусідні комплементарні пари повернені один відносно одного на 36 °. Діаметр спіралі 20Å, причому пуриновий нуклеотид займає 12Å, а Піримідинові - 8Å.
Молекула ДНК більш низькій вологості отримала назву А-форми. А-форма утворюється в умовах менш високою гідратації і при більш високому вмісті іонів Na + або К +. Ця ширша правоспіральная конформація має 11 пар азотистих основ на виток. Площині азотистих основ мають більш сильний нахил до осі спіралі, вони відхилені від нормалі до осі спіралі на 20 °. Звідси випливає наявність внутрішньої порожнечі діаметром 5Å. Відстань між сусідніми нуклеотидами складає 0,23 нм, довжина витка - 2,5 нм, діаметр спіралі - 2,3 нм.
Спочатку вважали, що А-форма ДНК менш важлива. Однак в подальшому з'ясувалося, що А-форма ДНК, також як і В-форма, має величезне біологічне значення. А-форму має спіраль РНК-ДНК в комплексі матриця-запал, а також спіраль РНК-РНК і шпилькові структури РНК (2'-гідроксильна група рибози не дозволяє молекулам РНК утворювати По-форму). А-форма ДНК виявлена в суперечках. Встановлено, що А-форма ДНК в 10 разів стійкіше до дії УФ-променів, ніж В-форма.
А-форму і В-форму називають канонічними формами ДНК.
Форми С-Етакож правоспіральние, їх освіту можна спостерігати тільки в спеціальних експериментах, і, мабуть, вони не існують in vivo. З-форма ДНК має структуру, подібну до В-ДНК. Число пар основ на виток становить 9,33, довжина витка спіралі дорівнює 3,1 нм. Пари підстав нахилені на кут 8 градусів щодо перпендикулярного положення до осі. Жолобки за розмірами близькі до желобкам В-ДНК. При цьому головний жолобок трохи дрібніше, а мінорний жолобок - глибше. В С-форму можуть переходити природні і синтетичні полінуклеотіди ДНК.
| Таблиця 1. Характеристика деяких типів структур ДНК | |||
| Тип спіралі | A | B | Z |
| крок спіралі | 0,32 нм | 3,38 нм | 4,46 нм |
| закрученность спіралі | права | права | ліва |
| Число пар основ на виток | 11 | 10 | 12 |
| Відстань між площинами підстав | 0,256 нм | 0,338 нм | 0,371 нм |
| Конформація гликозидной зв'язку | анти | анти | анти-С син-Г |
| Конформація фуранозной циклу | С3 "-енд | С2 "-енд | С3 "-енд-Г С2 "-енд-Ц |
| Ширина борозенки, малої / великої | 1,11 / 0,22 нм | 0,57 / 1,17 нм | 0,2 / 0,88 нм |
| Глибина борозенки, малої / великої | 0,26 / 1,30 нм | 0,82 / 0,85 нм | 1,38 / 0,37 нм |
| Діаметр спіралі | 2,3 нм | 2,0 нм | 1,8 нм |
Структурні елементи ДНК
(Неканонічні структури ДНК)
До структурних елементів ДНК можна віднести незвичайні структури, обмежені якимись спеціальними послідовностями:
|

Z-форма ДНКбула відкрита в 1979 році при вивченні гексануклеотіда d (CG) 3 -. Її відкрив професор Массачусетського технологічного інституту Олександр Річ з співробітниками. Z-форма стала одним з найважливіших структурних елементів ДНК в зв'язку з тим, що її освіту спостерігалося в ділянках ДНК, де пурини чергуються з піримідину (наприклад, 5'-ГЦГЦГЦ-3 '), або в повторах 5'-ЦГЦГЦГ-3' , що містять метильований цитозин. Істотною умовою освіти і стабілізації Z-ДНК було присутність в ній пуринових нуклеотидів в син-конформації, що чергуються з піримідинових основ в анти-конформації.
Природні молекули ДНК в основному існують в правій В-формі, якщо вони не містять послідовностей типу (ЦГ) n. Однак, якщо такі послідовності входять до складу ДНК, то ці ділянки при зміні іонної сили розчину або катіонів, що нейтралізують негативний заряд на фосфодіефірнимі каркасі, можуть переходити в Z-форму, при цьому інші ділянки ДНК в ланцюзі залишаються в класичній В-формі. Можливість такого переходу вказує на те, що два ланцюги в подвійної спіралі ДНК знаходяться в динамічному стані і можуть розкручуватися один щодо одного, переходячи з правої форми в ліву і навпаки. Біологічні наслідки такої лабільності, що допускає конформаційні перетворення структури ДНК поки не цілком зрозумілі. Вважають, що ділянки Z-ДНК відіграють певну роль в регуляції експресії деяких генів і беруть участь в генетичній рекомбінації.
Z-форма ДНК - це левозакрученной подвійна спіраль, в якій фосфодіефірнимі остов розташований зигзагоподібно уздовж осі молекули. Звідси і назва молекули (zigzag) -ДHK. Z-ДНК - найменш скручена (12 пар основ на виток) і найбільш тонка з відомих в природі. Відстань між сусідніми нуклеотидами складає 0,38 нм, довжина витка - 4,56 нм, діаметр Z-ДНК - 1,8 нм. Крім того, зовнішній вигляд цієї молекули ДНК відрізняється наявністю однієї борозенки.
Z-форма ДНК була виявлена в клітинах прокаріотів і еукаріотів. В даний час отримані антитіла, здатні відрізняти Z-форму від В-форми ДНК. Ці антитіла зв'язуються з певними ділянками гігантських хромосом клітин слинних залоз дрозофіли (Dr. melanogaster). За реакцією зв'язування легко стежити через незвичайного будови цих хромосом, у яких більш щільні ділянки (диски) контрастують з менш щільними (междіскамі). Ділянки Z-ДНК розташовані в междісках. З цього випливає, що Z-форма реально існує в природних умовах, хоча розміри індивідуальних ділянок Z-форми поки невідомі.

(Перевертні) - найбільш відомі і часто зустрічаються в ДНК послідовності підстав. Паліндромом називають слово або фразу, яке читається зліва направо і навпаки однаково. Прикладами таких слів або фраз є: курінь, КОЗАК, ПОТОП, А РОЗА ВПАЛА НА ЛАПУ Азор. У застосуванні до ділянок ДНК цей термін (паліндром) означає однакове чергування нуклеотидів уздовж ланцюга справа наліво і зліва направо (подібно буквах в слові "курінь" та ін.).
Паліндром характеризується наявністю інвертованих повторів послідовностей підстав мають симетрію другого порядку щодо двох ланцюгів ДНК. Такі послідовності, по цілком зрозумілих причин, є самокомплементарнимі і мають схильність до утворення шпилькових або хрестоподібних структур (рис.). Шпильки допомагають регуляторним білкам дізнаватися місце списування генетичного тексту ДНК хромосом.
У тих випадках, коли інвертований повтор присутній в одній і тій же ланцюга ДНК така послідовність називається дзеркальним повторенням. Дзеркальні повтори не володіють властивостями самокомплементарності і, тому не здатні до формування шпилькових або хрестоподібних структур. Послідовності такого типу виявлені практично у всіх великих молекулах ДНК і можуть включати від всього декількох пар підстав до декількох тисяч пар основ.
Присутність палиндромов у вигляді хрестоподібних структур в клітині не доведене, хоча деяку кількість хрестоподібних структур виявлено в умовах in vivo в клітинах E. coli. Наявність в складі РНК або одноланцюгових ДНК самокомплементарних послідовностей є основною причиною згортання в розчинах нуклеїнової ланцюга в певну просторову структуру, що відрізняється формуванням безлічі "шпильок".

Н-форма ДНК- це спіраль, яку утворюють три ланцюги ДНК - потрійна спіраль ДНК. Являє собою комплекс уотсон-криковських подвійної спіралі з третьої одноцепочечной ниткою ДНК, яка укладається в її великий жолобок, з утворенням так званої хугстіновской пари.
Освіта подібного триплекса відбувається в результаті складання подвійної спіралі ДНК таким чином, що половина її ділянки залишається у вигляді подвійної спіралі, а друга половина роз'єднується. При цьому одна з роз'єднаних спіралей утворює нову структуру з першою половиною подвійної спіралі - потрійну спіраль, а друга виявляється неструктурованою, у вигляді однониткових ділянки. Особливістю цього структурного переходу є різка залежність від рН середовища, протони якої стабілізують нову структуру. Через цю особливість нова структураотримала назву Н-форми ДНК, утворення якої було виявлено в сверхспіралізованних плазмидах, що містять гомопурін-гомопірімідіновие ділянки, що представляють собою дзеркальний повтор.
У подальших дослідженнях була встановлена можливість здійснення структурного переходу деяких гомопурін-гомопірімідінових двунітіевих полинуклеотидов з утворенням трехнітіевой структури, що містить:
- одну гомопуріновую і дві гомопірімідіновие нитки ( Py-Pu-Py триплекс) [Хугстіновское взаємодія].
Складові блоки Py-Pu-Py триплекса - канонічні ізоморфні CGC + і TAT тріади. Стабілізація триплекса вимагає протонирования тріади CGC +, тому ці триплекси залежать від рН розчину.
- одну гомопірімідіновую і дві гомопуріновие нитки ( Py-Pu-Pu триплекс) [Зворотне хугстіновское взаємодія].
Складові блоки Py-Pu-Pu триплекса - канонічні ізоморфні CGG і TAA тріад. Істотною властивістю Py-Pu-Pu триплексів є залежність їх стабільності від присутності двозарядних іонів, причому для стабілізації триплексів різній послідовності необхідні різні іони. Оскільки для утворення Py-Pu-Pu триплексів не потрібно протонирования входять до їх складу нуклеотидів, такі триплекси можуть існувати при нейтральних pH.
Прим .: пряме і зворотне хугстіновское взаємодія пояснюється симетрією 1-метілтіміна: поворот на 180 ° призводить до того, що місце атома О4 займає атом О2, при цьому система водневих зв'язків зберігається.
Відомі два види потрійних спіралей:
- паралельні потрійні спіралі, в яких полярність третьої ланцюга збігається з полярністю гомопуріновой ланцюга Уотсон-криковських дуплексу
- антипаралельні потрійні спіралі, в яких полярності третьої і гомопуріновой ланцюгів протилежні.

G-квадруплекс- 4-х спіральна ДНК. Така структура утворюється в разі, якщо є чотири гуаніну, які утворюють так званий G-квадруплекс - хоровод з чотирьох гуанін.
Перші натяки на можливість утворення таких структур були отримані задовго до проривної роботи Уотсона і Крика - ще в 1910 році. Тоді німецький хімік Івар Банг виявив, що один з компонентів ДНК - гуанозінових кислота - при високих концентраціях утворює гелі, в той час як інші складові частини ДНК такою властивістю не володіють.
У 1962 році за допомогою рентгеноструктурного методу вдалося встановити структуру осередку цього гелю. Вона виявилася складена з чотирьох залишків гуаніну, що зв'язують один одного по колу і утворюють характерний квадрат. У центрі зв'язок підтримує іон металу (Na, K, Mg). Такі ж структури можуть утворюватися і в ДНК, якщо в ній багато гуаніну. Ці плоскі квадрати (G-квартети) складаються в стопки, і виходять досить стійкі, щільні структури (G-квадруплекси).
У четирехспіральние комплекси можуть сплітаються чотири окремі ланцюжки ДНК, але це скоріше є винятком. Найчастіше єдина нитка нуклеїнової кислоти просто зав'язується у вузол, утворюючи характерні потовщення (наприклад, на кінцях хромосом), або двуцепочечной ДНК на якомусь багатому гуаніном ділянці утворює локальний квадруплекс.
Найбільш вивчено існування квадруплексов на кінцях хромосом - на теломери і в онкопромоторах. Однак до сих пір повне уявлення про локалізацію такої ДНК в людських хромосомах не відомо.
Всі ці незвичайні структури ДНК в лінійній формі нестабільні в порівнянні з В-формою ДНК. Однак ДНК часто існує в кільцевої формі топологічного напруги, коли у неї є так звана сверхспіралізаціі. У цих умовах легко утворюються неканонічні структури ДНК: Z-форми, "хрести" і "шпильки", H-форми, гуанінових квадруплекси і i-мотив.
- Суперспіралізірованная форма - наголошується при виділенні з ядра клітини без пошкодження пентозо-фосфатного остову. Має форму сверхскрученних замкнутих кілець. У сверхскрученном стані подвійна спіраль ДНК хоча б один раз "перекручена сама на себе", т. Е. Містить хоча б один супервіток (приймає форму вісімки).
- Релаксувати стан ДНК - спостерігається при одиночному розриві (розрив однієї нитки). При цьому супервитки зникають і ДНК приймає форму замкнутого кільця.
- Лінійна форма ДНК - спостерігається при розриві двох ниток подвійної спіралі.
Третинна структура ДНК
Третинна структура ДНКутворюється в результаті додаткового скручування в просторі двуспиральной молекули - її суперспіралізації. Суперспіралізації молекули ДНК в клітині на відміну від прокаріотів здійснюється у формі комплексів з білками.
ДНК еукаріот майже вся знаходиться в хромосомах ядер, лише невелика кількість її міститься в мітохондріях, а у рослин і в пластидах. Основна речовина хромосом еукаріотів (в тому числі і хромосом людини) - це хроматин, що складається з двухцепочечной ДНК, гістонових і негістонових білків.
Гістонові білки хроматину
Гістони - прості білки, складають до 50% хроматину. У всіх вивчених клітинах тварин і рослин виявлено п'ять основних класів гістонів: H1, H2A, H2B, H3, H4, що розрізняються за розмірами, амінокислотним складомі величиною заряду (завжди позитивний).
Гістон Н1 ссавців складається з одного поліпептидного ланцюга, що містить приблизно 215 амінокислот; розміри інших же гістонів варіюють від 100 до 135 амінокислот. Всі вони спіралізують і скручені в глобулу діаметром близько 2,5 нм, містять незвично велику кількість позитивно заряджених амінокислот лізину і аргініну. Гістони можуть бути ацетильованого, метиловані, фосфорильованій, поли (АДФ) -рібозіліровани, а гістони Н2А і Н2В - ковалентно пов'язані з убіквітин. Яка роль таких модифікацій в становленні структури і виконанні функцій гистонами до кінця поки не з'ясовано. Передбачається, що в цьому полягає їх здатність взаємодіяти з ДНК і забезпечувати один з механізмів регуляції дії генів.
Гістони взаємодіють з ДНК в основному через іонні зв'язку (сольові містки), що утворюються між негативно зарядженими фосфатними групами ДНК і позитивно зарядженими лізінових і аргініновая залишками гістонів.
Негістонові білки хроматину
Негістонові білки на відміну від гістонів дуже різноманітні. Виділено до 590 різних фракцій ДНК-зв'язуючих негістонових білків. Їх ще називають кислими білками, так як в їх структурі переважають кислі амінокислоти (вони є Поліаніонна). З різноманітністю негістонових білків пов'язують специфічну регуляцію активності хроматину. Наприклад ферменти, необхідні для реплікації та експресії ДНК, можуть зв'язуватися з хроматином тимчасово. Інші білки, скажімо, які беруть участь в різних процесах регуляції, зв'язуються з ДНК тільки в специфічних тканинах або на певних стадіях диференціації. Кожен білок комплементарен певній послідовності нуклеотидів ДНК (сайт ДНК). До цієї групи відносять:
- сімейство сайт-специфічних білків типу "цинкові пальці". Кожен "цинковий палець" дізнається певний сайт, що складається з 5 нуклеотидних пар.
- сімейство сайт-специфічних білків - гомодимера. Фрагмент такого білка, що контактує з ДНК, має структуру "спіраль-поворот-спіраль".
- білки високої рухливості (HMG-білки - від англ, high mobility gel proteins) - група структурних і регуляторних білків, які постійно асоційовані з хроматином. Вони мають молекулярну масу менше 30 кД і характеризуються високим вмістом заряджених амінокислот. Завдяки невеликій молекулярній масі HMG-білки мають високу рухливістю при електрофорезі в поліакриламідному гелі.
- ферменти реплікації, транскрипції і репарації.
За участю структурних, регуляторних білків і ферментів, які беруть участь в синтезі ДНК і РНК, нитка нуклеосом перетворюється в висококонденсований комплекс білків і ДНК. Освічена структура в 10 000 разів коротше вихідної молекули ДНК.

хроматин
Хроматин - це комплекс білків з ядерної ДНК і неорганічними речовинами. Основна частина хроматину неактивна. Вона містить щільно упаковану, конденсовану ДНК. Це гетерохроматин. Розрізняють конститутивним, генетично неактивний хроматин (сателітна ДНК) складається з неекспрессіруемих областей, і факультативний - неактивний в ряду поколінь, але при певних обставинах здатний еспрессіровать.
Активний хроматин (еухроматин) неконденсоване, тобто упакований менш щільно. У різних клітинах його зміст становить від 2 до 11%. У клітинах головного мозку його найбільше - 10-11%, в клітинах печінки - 3-4 і нирок - 2-3%. Відзначається активна транскрипція еухроматину. При цьому його структурна організація дозволяє використовувати одну й ту ж генетичну інформацію ДНК, властиву даному видуорганізму, по-різному в спеціалізованих клітинах.
В електронному мікроскопі зображення хроматину нагадує намисто: кулясті потовщення розміром близько 10 нм, розділені ниткоподібними перемичками. Ці кулясті потовщення названі нуклеосомами. Нуклеосома є структурною одиницею хроматину. Кожна нуклеосома містить сверхспіральний сегмент ДНК довжиною 146 пар нуклеотидів, намотаний з утворенням 1,75 лівих витків на нуклеосомної кор. Нуклеосомної кор - це гістонові октамер, що складається з гістонів Н2А, Н2В, Н3 і Н4, по дві молекули кожного виду (рис. 9), який виглядає як диск діаметром 11 нм і товщиною 5,7 нм. П'ятий гистон, Н1, не входить до складу нуклеосомної кора і не бере участі в процесі намотування ДНК на гістонові октамер. Він контактує з ДНК в тих місцях, де подвійна спіраль входить і виходить з нуклеосомної кора. Це межкоровие (лінкерних) ділянки ДНК, довжина яких варіює залежно від типу клітин від 40 до 50 нуклеотидних пар. В результаті цього варіює і довжина фрагмента ДНК, що входить до складу нуклеосом (від 186 до 196 нуклеотидних пар).
До складу нуклеосом входить приблизно 90% ДНК, інша її частина припадає на линкер. Вважається, що нуклеосоми - це фрагменти "мовчазного" хроматину, а линкер - активного. Однак нуклеосоми можуть розгортатися і переходити в лінійну форму. Розгорнуті нуклеосоми є вже активним хроматином. Так наочно проявляється залежність функції від структури. Можна вважати, що чим більше хроматину знаходиться в складі глобулярних нуклеосом, тим менше він активний. Очевидно, в різних клітинах неоднакова частка покоїться хроматину пов'язана з кількістю таких нуклеосом.
На електронно-мікроскопічних фотографіях в залежності від умов виділення і ступеня розтягування хроматин може виглядати не тільки як довга нитка з потовщеннями - "намистинками" нуклеосом, а й як більш коротка і більш щільна фібрила (волокно) діаметром 30 нм, утворення якої спостерігається при взаємодії гистона Н1, пов'язаного з лінкерних ділянкою ДНК і гистона Н3, що призводить до додаткового скручування спіралі з шести нуклеосом на виток з утворенням соленоїда діаметром 30 нм. При цьому гістонові білок може перешкоджати транскрипції ряду генів і таким чином регулювати їхню активність.
В результаті описаних вище взаємодій ДНК з гистонами сегмент подвійної спіралі ДНК з 186 пар основ із середнім діаметром 2 нм і довжиною 57 нм перетворюється в спіраль діаметром 10 нм і довжиною 5 нм. При подальшому стисненні цієї спіралі до волокна діаметром 30 нм ступінь конденсації збільшується ще в шість разів.
В кінцевому підсумку упаковка дуплексу ДНК з п'ятьма гистонами призводить до 50-кратної конденсації ДНК. Однак навіть така висока ступінь конденсації не може пояснити майже 50 000 - 100 000-кратне ущільнення ДНК в метафазної хромосомі. На жаль деталі подальшої упаковки хроматину аж до метафазної хромосоми поки не відомі, тому можна розглядати лише загальні особливості цього процесу.
Рівні компактизации ДНК в хромосомах
Кожна молекула ДНК упакована в окрему хромосому. У диплоїдних клітинах людини міститься 46 хромосом, які розташовуються в ядрі клітини. Загальна довжина ДНК всіх хромосом клітини становить 1,74 м, проте діаметр ядра, в яке упаковані хромосоми, в мільйони разів менше. Така компактна укладання ДНК в хромосомах і хромосом в ядрі клітини забезпечується різноманітними, гістонові і негістонових білками, взаємодіючими в певній послідовності з ДНК (див вище). Компактизація ДНК в хромосомах дозволяє зменшити її лінійні розміри приблизно в 10 000 разів - умовно з 5 см до 5 мкм. Виділяють кілька рівнів компактизации (рис. 10).


- подвійна спіраль ДНК - негативно заряджена молекула діаметром 2 нм і довжиною кілька сантиметрів.
- нуклеосомної рівень- хроматин виглядає в електронному мікроскопі як ланцюжок "намистин" - нуклеосом - "на нитки". Нуклеосома - це універсальна структурна одиниця, яка виявляється як у еухроматину, так і в гетерохроматин, в інтерфазних ядрі і метафазних хромосомах.
Нуклеосомної рівень компактизації забезпечується спеціальними білками - гистонами. Вісім позитивно заряджених гістонових доменів утворюють кор (серцевину) нуклеосоми на яку намотується негативно заряджена молекула ДНК. Це дає вкорочення в 7 разів, при цьому діаметр збільшується з 2 до 11 нм.
- соленоїдний рівень
Соленоїдний рівень організації хромосом характеризується скручуванням нуклеосомної нитки і освітою з неї більш товстих фібрил 20-35 нм в діаметрі - соленоїдів або супербідов. Крок соленоїда дорівнює 11 нм, на один виток припадає близько 6-10 нуклеосом. Соленоидная упаковка вважається найбільш імовірною, ніж супербідная, згідно з якою фібрила хроматину діаметром 20-35 нм представляє собою ланцюг гранул, або супербідов, кожна з яких складається з восьми нуклеосом. На соленоїдного рівні лінійний розмір ДНК скорочується в 6-10 разів, діаметр збільшується до 30 нм.
- петлевий рівень
Петлевой рівень забезпечується негістонових сайт-специфічними ДНК-зв'язуючими білками, які розпізнають певні послідовності ДНК і зв'язуються з ними, утворюючи петлі приблизно по 30-300 тисяч пар основ. Петля забезпечує експресію генів, тобто петля є не тільки структурним, але і функціональним утворенням. Скорочення на цьому рівні відбувається в 20-30 разів. Діаметр збільшується до 300 нм. Петлеподібні структури типу "лампових щіток" в ооцитах земноводних можна бачити на цитологічних препаратах. Ці петлі, мабуть, суперспіралізовани і являють собою домени ДНК, що відповідають, ймовірно, одиницям транскрипції і реплікації хроматину. Специфічні білки фіксують підстави петель і, можливо, деякі їхні внутрішні ділянки. Петлеподібна доменна організація сприяє укладанню хроматину в метафазних хромосомах в спіральні структури більш високих порядків.
- доменний рівень
Доменний рівень організації хромосом вивчений недостатньо. На даному рівні відзначається утворення петльових доменів - структур з ниток (фібрил) товщиною 25-30 нм, які містять 60% білка, 35% ДНК і 5% РНК, практично не видно у всіх фазах клітинного циклу за винятком мітозу і кілька безладно розподілені по клітинного ядра. Петлеподібні структури типу "лампових щіток" в ооцитах земноводних можна бачити на цитологічних препаратах.
Петльові домени своїм підставою прикріплюються до внутрішньоядерних білкового матриксу в так званих вбудованих місцях прикріплення, часто позначаються як MAR / SAR-послідовності (MAR, від англ. Matrix associated region; SAR, від англ. Scaffold attachment regions) - фрагментах ДНК довжиною в кілька сотень пар основ, які характеризуються високим вмістом (> 65%) А / Т пар нуклеотидів. Кожен домен, мабуть, має одну точку початку реплікації і функціонує як автономна сверхспіральная одиниця. Будь-петельний домен містить безліч одиниць транскрипції, функціонування яких, ймовірно, координується - весь домен перебувати або в активному, або в неактивному стані.
На доменному рівні в результаті послідовної упаковки хроматину присходит зменшення лінійних розмірів ДНК приблизно в 200 разів (700 нм).
- хромосомний рівень
На хромосомному рівні відбувається конденсація профазних хромосоми в метафазну з ущільненням петельних доменів навколо осьового каркаса негістонових білків. Ця суперспіралізації супроводжується фосфорилюванням в клітці всіх молекул H1. В результаті метафазну хромосому можна зобразити у вигляді щільно укладених соленоїдних петель, згорнутих в тугу спіраль. Типова хромосома людини може містити до 2600 петель. Товщина такої структури досягає 1400 нм, (дві хроматиди), а молекула ДНК при цьому коротшає в 104 разів, тобто з 5 см розтягнутої ДНК до 5 мкм.
функції хромосом
У взаємодії з позахромосомних механізмами хромосоми забезпечують
- зберігання спадкової інформації
- використання цієї інформації для створення і підтримки клітинної організації
- регуляцію зчитування спадкової інформації
- самоудвоение генетичного матеріалу
- передачу генетичного матеріалу від материнської клітини дочірнім.
Існують дані, що при активації ділянки хроматину, тобто при транскрипції, з нього можна зупинити видаляються спочатку гистон H1, а потім і октет гістонів. Це викликає деконденсація хроматину, послідовний перехід 30-нанометровій фібрили хроматину в 10-нанометровій нитку і її подальше розгортання в ділянки вільної ДНК, тобто втрату нуклеосомної структури.
Екологія пізнання. Наука і відкриття: Що таке життя? Це питання стало рушійною силою розвитку генетики (від грецького genetikos - «що відноситься до народження, походженням») - науки про походження життя, в центрі уваги якої ось вже більше 50 років перебуває молекула ДНК.
Що таке життя? Це питання стало рушійною силою розвитку генетики (від грецького genetikos - «що відноситься до народження, походженням») - науки про походження життя, в центрі уваги якої ось вже більше 50 років перебуває молекула ДНК.
Відкриття, яке перевернуло світ
«Ми тільки що відкрили секрет життя!» - так 28 лютого 1953 року Френсіс Крік і Джеймс Уотсон повідомили про своє відкриття структури ДНК. Що нового привнесло воно в науки про життя? До цього було відомо, що ДНК - велика молекула, в якій за допомогою «чотирибуквене алфавіту» записана інформація про будову і властивості живих істот. Але залишалося незрозумілим, як ця інформація передається з покоління в покоління і матеріалізується в ці самі структури і властивості, а також яка просторова структура ДНК.
Розгадка структури ДНК допомогла вченим зрозуміти механізми її копіювання та матеріалізації. ДНК складається з двох ланцюгів, які комплементарні (додатковими) один одному. Копіювання ДНК відбувається за рахунок добудови на кожній вихідної ланцюга ДНК, як на матриці, додаткової до неї копії. Так з однієї подвійної спіралі ДНК виходять дві абсолютно ідентичні їй подвійніспіралі, що і необхідно для збереження генетичної інформації при розподілі материнської клітини на дві дочірні. Матричний принцип лежить також в основі поетапної матеріалізації генетичної інформації: на одному з ланцюжків ДНК утворюється комплементарная їй ланцюг іншої інформаційної молекули - РНК, яка, в свою чергу, служить матрицею для синтезу білків, від кількості та якості яких залежать структури і властивості конкретного організму.
Наскільки це відкриття значимо для осягнення таємниці життя? З одного боку, знання структури ДНК явно недостатньо для того, щоб відповісти на питання «що таке життя?». Але з іншого - саме це відкриття зробило «науковим» дуже древній і дуже важливе питанняпро взаємозв'язок потенційного і проявленого - на прикладі зв'язку інформації про структури і властивості організму з самими структурами і властивостями. І не тільки поставило це питання, але і дало ключ до відповіді на нього. Цей ключ - матричний принцип, принцип комплементарності.
Шлях від гена до ознаки
Що означає класична фраза з підручника: «ДНК - носій генетичної інформації»? Як генетична інформація пов'язана зі структурою ДНК? Яким чином інформація втілюється в конкретних властивостях організму? Якщо за точку відліку генетичної інформації прийняти структуру ДНК і далі слідувати структурної моделі, то шлях від гена до ознаки буде виглядати так: в послідовності ДНК зашифровані всі властивості організму; лінійна структура конкретного гена однозначно визначає лінійну структуру відповідного йому білка, яка, в свою чергу, однозначно визначає роль цього білка в формуванні тієї чи іншої ознаки.
Іншими словами, «ДНК народжує РНК; РНК народжує білок, а білок народжує нас з вами »(Ф. Крик). Якщо це вірно, то для того, щоб змінити ту чи іншу ознаку (наприклад, вилікувати хворобу, що має генетичне коріння), досить встановити відповідну йому послідовність ділянки ДНК і виправити її.
Але так чи так все просто? Чи достатньо знань (хоча вони безперечно вірні і необхідні!) Про структурні відповідностях на шляху від гена до ознаки, для того щоб зрозуміти і відтворити цей шлях?
Останні досягнення генетики показали, що недостатні. У 2003 році в рамках проекту «Геном людини» була повністю визначена лінійна структура ДНК людини (і багатьох інших простих і складних організмів). Як сказав один з учених, «як прочитані всі букви, якими написана товста книга, тепер ще зрозуміти б слова та їх зміст».
З'ясувалося, що власне генів (ділянок ДНК, що кодують білки) у людини близько 30 000, і це лише 1-3% всієї ДНК! Стільки ж генів у рослини Arabidopsis taliana і риби фугу. Більш того, 99% генів людини збігаються з генами миші, тобто людина має всього 300 генів, яких немає у миші. (Важко уявити, що у нас і мишей однакові ще і 99% ознак!)
Далі більше. Виявилося, що однозначна взаємозв'язок між геном і білком існує тільки у бактерій. А у людини можливе утворення багатьох білків на підставі одного гена (максимально відоме сьогодні число різних білків, що кодуються одним геном, - 40 000!) І виникнення багатьох функцій у одного білка. Виходить, що шлях від потенційного до проявленій, від генетичної інформації до ознакою аж ніяк не лінійний; що кожен ознака є результатом складних взаємодій багатьох генів і їх продуктів-білків; що саме поняття «ген» через свою неоднозначність навряд чи може служити «відправним пунктом» цього шляху.
взаємодія структур
Тіло людини складається з 10 в 14-го ступеня клітин. Всі вони мають абсолютно однакову ДНК, але істотно розрізняються за формою, розмірами і своїм завданням. Дозвіл цього парадоксу - у виборчому зчитуванні генетичної інформації. У кожній клітині активними є тільки ті гени, які їй в даний момент необхідні.
Вибірковість забезпечують спеціальні гени-регулятори, які дозволяють або забороняють зчитування інформації з тієї чи іншої ділянки ДНК. Активність гена залежить і від його оточення в просторі клітинного ядра. Зміна оточення, викликана переміщенням самого гена або когось із його сусідів, здатна змінити його активність ( «вимкнути» або «включити» ген).
Наприклад, в геномі людини є маса потенційно небезпечних вірусних генів і протоонкогенов (здатних викликати ракове переродження клітини). Вони можуть тривалий час (і все життя) поводитися цілком мирно і навіть працювати на благо клітини, до тих пір поки переміщення їх самих або когось з оточення не виявить в цих генах агресивні потенціали. На щастя, можуть відбутися інші переміщення, які вгамують «бунтівника» або включать захисні механізми.
Отже, носій генетичної інформації перемістився з рівня гена (конкретної ділянки ДНК) на епігенетичні (від латинського «над», «понад») рівень взаємодії генів між собою і з іншими структурами ядра клітини (99% Неген ДНК і білками). Припустимо, наука розшифрує механізм цієї взаємодії. Чи призведе це до розкриття таємниці життя? Життя - це тільки лише структура? А якщо ні, чи варто в пошуках розгадки таємниці життя обмежуватися взаємодією структур?
Хто стереже сторожа?
Як з єдиної клітини в результаті 46 поділів виходить не безформна маса з 1014 клітин, а дуже характерне тіло кожного з нас? Послідовно подвоюючись, клітини не тільки самі стають різними, але ще і формують різні частини тіла в потрібний часі в потрібному місці. Що керує організацією клітин в часі і просторі?
Ціле, яке якісно більше простої суми складових його частин-клітин. І це не суперечить тому, що організм утворюється з однієї клітини, - питання в тому, що для цієї клітини втілює «волю цілого». Пошуки подібного упорядочивающего фактора вилилися на початку XX століття в концепцію морфогенетичного поля. Її основоположником став російський вчений А.Г. Гурвич. Коли Гурвич працював над теорією поля, молекула ДНК вважалася складовою частиною хромосом, і їй не надавали особливого значення.
У 1944 році вчений опублікував свою працю «Теорія біологічного поля». Цей рік став доленосним для всієї генетики, визначивши шлях її розвитку на кілька десятиліть вперед. У центрі уваги вчених виявилася молекула ДНК, оскільки було доведено, що саме їй належить провідна роль у передачі спадкової інформації. Не за горами був і 1953-й ... У результаті всю свою увагу наука зосередила на структурі ДНК, яку фактично стала ототожнювати з яку переносять нею інформацією, а теорія біологічного поля виявилася не в пошані. Але дослідження в цій області тривали, і всі ці роки два шляхи пізнання таємниці життя йшли паралельно ...
Перехід в нове тисячоліття змінив співвідношення сил в науках про життя. Все більше вчених приходять до того, що структурний ключ в пізнанні живого необхідний, але недостатній; що різні підходи не виключають один одного, а утворюють об'єднаний шлях наукового пошуку; що по суті своїй структурний підхід і теорія поля комплементарні.
Згадаймо: саме припущення про комплементарності ланцюгів ДНК стало ключем до розшифровки її структури, а саме відкриття 1953 року виявився можливим завдяки комплементарності зусиль представників різних галузей науки - фізиків, хіміків, біологів. Може бути, об'єднана наука нового тисячоліття не тільки остаточно примирить різні наукові підходи (наприклад, структурний і польовий), але і звернеться до плодів «ненаукового» шляху пізнання таємниці життя - тисячолітньої мудрості людства, - «ненаукового», оскільки цей шлях йде корінням в ті часи, коли науки не було і в помині.
Звернення до джерел давньої мудрості здатне дати науці ключі від дверей, за якими прихована таємниця. Але щоб це сталося, настільки різні шляхипізнання повинні десь «перетнутися». Одним з таких «перехресть» може стати концепція формотворного поля (біологічного, морфогенетичного, інформаційного), яка виросла на грунті сучасної науки і висхідна до явищ, які розглядали в древніх джерелах.
Останні кажуть про те, що людина складається з декількох тіл, або принципів, які не є окремими, незалежними частинами, а взаємопроникають і взаімоформіруют один одного; що видиме, щільне, фізичне тіло є провідником, носієм більш тонкоматеріальних тіл, які з його допомогою виявляються в фізичному світі і взаємодіють з ним; що «складанням» - формуванням фізичного тіла з елементів фізичної матерії - управляє саме «щільне» з цих тіл, астральне тіло-прообраз (план, матриця).
Сучасна наука знає, що кожна знову утворилася клітина бере участь у формуванні організму згідно з індивідуальною «інструкції» (активні, або включені, гени) і що у батька і у нащадків цієї клітини можуть бути зовсім інші «інструкції». Але що і як погодить перемикання індивідуальних «програм» розвитку мільярдів клітин, поки не ясно.
Теорія біологічного поля передбачає, що узгодження є функція цілого, яке і є тим самим полем, матрицею або моделлю; що кожна знову утворилася клітина за допомогою власного генетичного апарату підключається до єдиного «плану» розвитку організму, отримує звідти індивідуальні «інструкції» і реалізує їх в рамках власної програми поведінки.
Виходить, що генетичний апарат клітини складається, як мінімум, із трьох функціональних блоків: сприймає «антени», «пульта управління» активністю генів і «виконавчої» частини - генів, відповідальних за освіту конкретних білків. Згадаймо, що на частку генів припадає лише 1-2% від всієї клітинної ДНК. У решти 98-99% ДНК вже виявлені структури, що відносяться до другого «керуючому блоку». А що грає роль «антени»? Де відбувається «зустріч двох світів» - інформаційного поля і генетичних структур, що втілюють цю інформацію в конкретну фізичну тіло?
Чому б не припустити, що роль антени, здатної вловлювати, трансформувати і передавати сигнали хвильової природи, теж грає ДНК? До цього розташовує і спіральна структура «молекули життя» (багато технічних антени мають форму спіралі), і такі її властивості, як здатність проводити електричний струм, можливість резонансного збудження поздовжніх коливань під дією радіохвиль, а також здатність до лазерної генерації світла після попередньої «накачування ».
Якщо ДНК може працювати на прийом інформації, що забезпечує життєву активність клітин, то їй зовсім не обов'язково постійно зберігати цю інформацію в своїй структурі. Як, наприклад, мозку людини, щоб успішно керувати системами життєзабезпечення організму, не обов'язково бути «вмістилищем» розуму, а досить грати роль посередника між свідомістю і тілом: він сприймає інформацію з плану свідомості і «переводить» її на мову управління тілом.
І ясно, чому в разі пошкоджень структури ДНК (або мозкових структур) страждає фізичне тіло. Адже всім відомо, що при несправності в телевізорі хоча б однієї деталі зображення на його екрані сильно спотворюється, а якщо телевізор позбавити антени або вимкнути його з мережі, на екрані взагалі нічого не з'явиться.
ДНК - сполучна ланка між «моделлю» фізичного тіла і її конкретним втіленням. Мозок - посередник між розумом і тілом. Розум пов'язує життя і форму її прояву і дозволяє життя, укладеної в формі, пізнавати саму себе. За допомогою цього чудового інструменту людина має можливість вивчати навколишній світ і знаходити в ньому ключі до пізнання свого внутрішнього світу. Так народжується об'єднаний шлях, що веде до пізнання таємниці життя. Бо людина є найбільша з таємниць - таємниця взаємозв'язку землі і неба.опубліковано
P.S. І пам'ятайте, всього лише змінюючи своє споживання - ми разом змінюємо світ! © econet
Приєднуйтесь до нас в
Після відкриття принципу молекулярної організації такого речовини, як ДНК в 1953 році, почала розвиватися молекулярна біологія. Далі в процесі досліджень вчені з'ясували як рекомбеніруется ДНК, її склад і як влаштований наш людський геном.
Кожен день на молекулярному рівні відбуваються складні процеси. Як влаштована молекула ДНК, з чого вона складається? І яку роль відіграють в клітці молекули ДНК? Розповімо докладно про всі процеси, що відбуваються всередині подвійного ланцюга.
Що таке спадкова інформація?
Отже, з чого все починалося? Ще в 1868 знайшли в ядрах бактерій. А в 1928 р Н. Кольцов висунув теорію про те, що саме в ДНК зашифрована вся генетична інформація про живий організм. Потім Дж. Уотсон і Ф. Крик знайшли модель всім тепер відомої спіралі ДНК в 1953 році, за що заслужено отримали визнання і нагороду - Нобелівську премію.
Що таке взагалі ДНК? Ця речовина складається з 2 об'єднаних ниток, точніше спіралей. Ділянка такого ланцюжка з певною інформацією називається геном.

У ДНК зберігається вся інформація про те, що за білки будуть формуватися і в якому порядку. Макромолекула ДНК - це матеріальний носій неймовірно об'ємної інформації, яка записана суворої послідовністю окремих цеглинок - нуклеотидів. Всього нуклеотидів 4, вони доповнюють один одного хімічно і геометрично. Цей принцип доповнення, або комплементарності, в науці буде описаний пізніше. Це правило грає ключову роль в кодуванні і декодуванні генетичної інформації.
Так як нитка ДНК неймовірно довга, повторень в цій послідовності не буває. У кожної живої істоти власна унікальна ланцюжок ДНК.
функції ДНК
До функцій відносяться зберігання спадкової інформації та її передача потомству. Без цієї функції геном виду не міг би зберігатися і розвиватися протягом тисячоліть. Організми, які зазнали серйозних мутації генів, частіше не виживають або втрачають здатність виробляти потомство. Так відбувається природний захист від виродження виду.

Ще одна істотно важлива функція - реалізація інформації, що зберігається. Клітка не може створити жодного життєво важливого білка без тих інструкцій, які зберігаються в подвійній ланцюжку.
Склад нуклеїнових кислот
Зараз вже достеменно відомо, з чого складаються самі нуклеотиди - цеглинки ДНК. До їх складу входять 3 речовини:
- Ортофосфорна кислота.
- Азотистих основ. Піримідинові підстави - які мають тільки одне кільце. До них відносять тимін і цитозин. Пуринові основи, в складі яких присутні 2 кільця. Це гуанін і аденін.
- Сахароза. У складі ДНК - дезоксирибоза, В РНК - рибоза.
Число нуклеотидів завжди дорівнює числу азотистих основ. У спеціальних лабораторіях розщеплюють нуклеотид і виділяють з нього азотистих основ. Так вивчають окремі властивості цих нуклеотидів і можливі мутації в них.
Рівні організації спадкової інформації
Розділяють 3 рівня організації: генний, хромосомний і геномної. Вся інформація, необхідна для синтезу нового білка, міститься на невеликій ділянці ланцюжка - гені. Тобто ген вважається нижчий і найпростіший рівень кодування інформації.

Гени, в свою чергу, зібрані в хромосоми. Завдяки такій організації носія спадкового матеріалу групи ознак за певними законами чергуються і передаються від одного покоління до іншого. Треба зауважити, генів в організмі неймовірно багато, але інформація не губиться, навіть коли багато разів рекомбеніруется.
Розділяють кілька видів генів:
- за функціональним призначенням виділяють 2 типу: структурні і регуляторні послідовності;
- за впливом на процеси, що протікають в клітині, розрізняють: супервітальние, летальні, умовно летальні гени, а також гени мутатори і антімутатори.
Розташовуються гени уздовж хромосоми в лінійному порядку. У хромосомах інформація сфокусована НЕ врозкид, існує певний порядок. Існує навіть карта, в якій відображені позиції, або локуси генів. Наприклад, відомо, що в хромосомі № 18 зашифровані дані про колір очей дитини.
А що ж таке геном? Так називають всю сукупність нуклеотиднихпослідовностей в клітині організму. Геном характеризує цілий вид, а не окрему особина.
Який генетичний код людини?
Справа в тому, що весь величезний потенціал людського розвиткузакладений вже в період зачаття. Вся спадкова інформація, яка необхідна для розвитку зиготи і зростання дитини вже після народження, зашифрована в генах. Ділянки ДНК і є самі основні носії спадкової інформації.
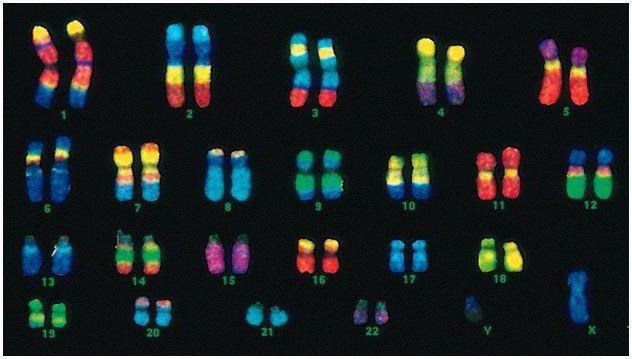
У людини 46 хромосом, або 22 соматичні пари плюс по одній визначальною підлогу хромосомі від кожного з батьків. Цей диплоїдний набір хромосом кодує весь фізичний вигляд людини, його розумові та фізичні здібності і схильність до захворювань. соматичні хромосомизовні невиразні, але несуть вони різну інформацію, так як одна з них від батька, інша - від матері.
Чоловік код відрізняється від жіночого останньою парою хромосом - ХУ. Жіночий диплоїдний набір - це остання пара, ХХ. Чоловікам дістається одна Х-хромосома від біологічної матері, і потім вона передається дочкам. Статева У-хромосома передається синам.
Хромосоми людини значно різняться за розміром. Наприклад, найменша пара хромосом - №17. А найбільша пара - 1 і 3.
Діаметр подвійної спіралі у людини - всього 2 нм. ДНК настільки щільно закручена, що вміщається в маленькому ядрі клітини, хоча її довжина сягатиме 2 метрів, якщо її розкрутити. Довжина спіралі - це сотні мільйонів нуклеотидів.
Як передається генетичний код?
Отже, яку роль відіграють в клітці молекули ДНК при діленні? Гени - носії спадкової інформації - знаходяться всередині кожної клітини організму. Щоб передати свій код дочірньому організму, багато істот ділять своє ДНК на 2 однакові спіралі. Це називається реплікацією. У процесі реплікації ДНК розплітається і спеціальні «машини» доповнюють кожну ланцюжок. Після того як подвоїться генетична спіраль, починає ділитися ядро і всі органели, а потім і вся клітина.
Але у людини інший процес передачі генів - статевий. Ознаки батька і матері перемішуються, в новому генетичному коді міститься інформація від обох батьків.
Зберігання та передача спадкової інформації можливі завдяки складній організації спіралі ДНК. Адже як ми говорили, структура білків зашифрована саме в генах. Раз створити під час зачаття, цей код на протязі всього життя буде копіювати сам себе. Каріотип (особистий набір хромосом) не змінюється під час відновлення клітин органів. Передача ж інформації здійснюється за допомогою статевих гамет - чоловічих і жіночих.
Передавати свою інформацію потомству не здатні тільки віруси, що містять один ланцюжок РНК. Тому, щоб відтворюватися, їм потрібні клітини людини або тварини.
Реалізація спадкової інформації
У ядрі клітини постійно відбуваються важливі процеси. Вся інформація, записана в хромосомах, використовується для побудови білків з амінокислот. Але ланцюжок ДНК ніколи не покидає ядро, тому тут потрібна допомога іншого важливого з'єднання = РНК. Якраз РНК здатна проникнути через мембрану ядра і взаємодіяти з ланцюжком ДНК.
За допомогою взаємодії ДНК і 3 видів РНК відбувається реалізація всієї закодованої інформації. На якому рівні відбувається реалізація спадкової інформації? Всі взаємодії відбуваються на рівні нуклеотидів. Інформаційна РНК копіює ділянку ланцюга ДНК і приносить цю копію в рибосому. Тут починається синтез з нуклеотидів нової молекули.
Для того щоб іРНК могла скопіювати важливу частину ланцюга, спіраль розгортається, а потім, після закінчення процесу перекодування, знову відновлюється. Причому цей процес може відбуватися одночасно на 2 сторонах 1 хромосоми.
принцип комплементарності
Складаються з 4 нуклеотидів - це аденін (А), гуанін (G), цитозин (С), тимін (T). Сполучені вони водневими зв'язками за правилом комплементарності. Роботи Е. Чаргаффа допомогли встановити це правило, тому що вчений помітив деякі закономірності в поведінці цих речовин. Е. Чаргафф відкрив, що молярне відношення аденіну до тимін дорівнює одиниці. І точно так само ставлення гуаніну до цитозину завжди дорівнює одиниці.
На основі його робіт генетики сформували правило взаємодії нуклеотидів. Правило комплементарності говорить, що аденін з'єднується тільки з тиміном, а гуанін - з цитозином. Під час декодування спіралі і синтезу нового білка в рибосомі таке правило чергування допомагає швидко знайти необхідну амінокислоту, Яка прикріплена до транспортної РНК.
РНК і його види
Що таке спадкова інформація? нуклеотидів в подвійній ланцюга ДНК. А що таке РНК? У чому полягає її робота? РНК, або рибонуклеїнова кислота, допомагає витягувати інформацію з ДНК, декодувати її і на основі принципу комплементарності створювати необхідні клітинам білки.
Всього виділяють 3 види РНК. Кожна з них виконує строго свою функцію.
- Інформаційна (іРНК), Або ще її називають матрична. Вона заходить прямо в центр клітини, в ядро. Знаходить в одній з хромосом необхідний генетичний матеріал для побудови білка і копіює одну зі сторін подвійного ланцюга. Копіювання відбувається знову за принципом комплементарності.
- транспортна- це невелика молекула, у якої на одній стороні декодери-нуклеотиди, а на іншій стороні відповідні основному коду амінокислоти. Завдання тРНК - доставити в «цех», тобто в рибосому, де синтезує необхідну амінокислоту.
- рРНК - рибосомная.Вона контролює кількість білка, який продукується. Складається з 2 частин - амінокислотного і пептидного ділянки.
Єдина відмінність при декодуванні - у РНК немає тиміну. Замість тиміну тут присутній урацил. Але потім, в процесі синтезу білка, при тРНК все одно правильно встановлює всі амінокислоти. Якщо ж відбуваються якісь збої в декодуванні інформації, то виникає мутація.
Репарація пошкодженої молекули ДНК
Процес відновлення пошкодженої подвійний ланцюжка називається репарацією. В процесі репарації пошкоджені гени видаляються.

Потім необхідна послідовність елементів в точності відтворюватися і врізається назад в той же місце на ланцюгу, звідки було вилучено. Все це відбувається завдяки спеціальним хімічних речовин- ферментам.
Чому відбуваються мутації?
Чому деякі гени починають мутувати і перестають виконувати свою функцію - зберігання життєво необхідної спадкової інформації? Це відбувається через помилки при декодуванні. Наприклад, якщо аденін випадково замінений на тимін.
Існують також хромосомні і геномні мутації. Хромосомні мутації трапляються, якщо ділянки спадкової інформації випадають, подвоюються або взагалі переносяться і вбудовуються в іншу хромосому.

Геномні мутації найбільш серйозні. Їх причина - це зміна числа хромосом. Тобто коли замість пари - диплоїдного набору присутній в каріотипі тріплоідний набір.
Найбільш відомий приклад триплоїдного мутації - це синдром Дауна, при якому особистий набір хромосом 47. У таких дітей утворюється 3 хромосоми на місці 21-ї пари.
Відома також така мутація, як полиплодия. Але полиплодия зустрічається тільки у рослин.
Молекула ДНК складається з двох ниток, що утворюють подвійну спіраль. Вперше її структура була розшифрована Френсісом Криком і Джеймсом Уотсоном в 1953 році.
Спочатку молекула ДНК, що складається з пари нуклеотидних, закручених один навколо одного ланцюжків, породжувала питання про те, чому саме таку форму вона має. Вчені назвали цей феномен комплементарностью, що означає, що в її нитках один навпроти одного можуть перебувати виключно певні нуклеотиди. Наприклад, навпаки тиміну завжди варто аденін, а навпаки цитозину - гуанін. Ці нуклеотиди молекули ДНК і називаються комплементарними.
Схематично це зображується так:
Т - А
Ц - Г
Дані пари утворюють хімічну нуклеотидную зв'язок, яка визначає порядок розстановки амінокислот. У першому випадку вона трохи слабше. Зв'язок між Ц і Г міцніша. Некомплементарни нуклеотиди між собою пари не утворюють.
Про будову
Отже, будова молекули ДНК особливе. Таку форму вона має неспроста: справа в тому, що кількість нуклеотидів дуже велике, і для розміщення довгих ланцюжків необхідно багато місця. Саме з цієї причини ланцюжках притаманне спіральне закручування. Це явище названо спирализация, воно дозволяє ниткам зменшуватися десь в п'ять-шість разів.
Деякі молекули такого плану організм використовує дуже активно, інші - рідко. Останні, крім спирализации, піддаються ще й такий «компактній упаковці», як суперспіралізації. І тоді довжина молекули ДНК зменшується в 25-30 разів.

Що таке «упаковка» молекули?
В процесі суперспіралізації задіюються гістонові білки. Вони мають структуру і вид котушки для ниток або стрижня. На них і намотуються спіраль нитки, які стають відразу «компактно упакованими» і займають мало місця. Коли виникає необхідність використання тієї чи іншої нитки, вона змотується з котушки, наприклад, гістонові білка, і спіраль розкручується в дві паралельні ланцюжки. Коли молекула ДНК перебуває саме в такому стані, з неї можна зчитувати необхідні генетичні дані. Однак є одна умова. Отримання інформації можливо, тільки якщо структура молекули ДНК має розкручений вид. Хромосоми, доступні для зчитування, називаються еухроматин, а якщо вони суперсіпіралізовани, то це вже гетерохроматином.
нуклеїнові кислоти
Нуклеїнові кислоти, як і білки, є биополимерами. Головна функція - це зберігання, реалізація та передача спадкової (генетичної інформації). Вони бувають двох типів: ДНК і РНК (дезоксирибонуклеїнової і РНК). Мономерами в них виступають нуклеотиди, кожен з яких має в своєму складі залишок фосфорної кислоти, п'ятивуглецевий цукор (дезоксирибоза / рибоза) і азотна основа. У ДНК код входить 4 види нуклеотидів - аденін (А) / гуанін (Г) / цитозин (Ц) / тимін (Т). Вони відрізняються по міститься в їх складі азотистій підставі.
У молекулі ДНК кількість нуклеотидів може бути величезним - від декількох тисяч до десятків і сотень мільйонів. Розглянути такі гігантські молекули можна через електронний мікроскоп. У цьому випадку вдасться побачити подвійну ланцюг з полінуклеотидних ниток, які з'єднані між собою водневими зв'язками азотистих основ нуклеотидів.

дослідження
В ході досліджень вчені виявили, що види молекул ДНК у різних живих організмів відрізняються. Також було встановлено, що гуанін одного ланцюга може зв'язуватися тільки з цитозином, а тимін - з аденін. Розташування нуклеотидів одного ланцюга строго відповідає паралельній. Завдяки такій комплементарності полинуклеотидов молекула ДНК здатна до подвоєння і самовідтворення. Але спочатку комплементарні ланцюга під впливом спеціальних ферментів, що руйнують парні нуклеотиди, розходяться, а потім в кожній з них починається синтез якої бракує ланцюга. Це відбувається за рахунок наявних в великій кількостів кожній клітині вільних нуклеотидів. В результаті цього замість «материнської молекули» формуються дві «дочірні», ідентичні за складом і структурі, і ДНК-код стає вихідним. Даний процес є попередником клітинного ділення. Він забезпечує передачу всіх спадкових даних від материнських клітин дочірнім, а також всім наступним поколінням.
Як читається генний код?
Сьогодні обчислюється не тільки маса молекули ДНК - можна дізнатися і більш складні, раніше не доступні вченим дані. Наприклад, можна прочитати інформацію про те, як організм використовує власну клітку. Звичайно, спочатку відомості ці знаходяться в закодованому вигляді та мають вигляд якоїсь матриці, а тому її необхідно транспортувати на спеціальний носій, яким виступає РНК. Рибонуклеїнової кислоти під силу проникати в клітку через мембрану ядра і вже всередині зчитувати закодовану інформацію. Таким чином, РНК - це переносник прихованих даних з ядра в клітку, і відрізняється вона від ДНК тим, що в її склад замість дезоксирибози входить рибоза, а замість тиміну - урацил. Крім того, РНК одноцепочной.

синтез РНК
Глибокий аналіз ДНК показав, що після того як РНК покидає ядро, вона потрапляє в цитоплазму, де і може бути вбудована як матриця в рибосоми (спеціальні ферментні системи). Керуючись отриманою інформацією, вони можуть синтезувати відповідну послідовність білкових амінокислот. Про те, яку саме різновид органічної сполуки необхідно приєднати до формується білкової ланцюга, рибосома дізнається з триплетного коду. Кожній амінокислоті відповідає свій певний триплет, який її і кодує.
Після того як формування ланцюжка завершено, вона набуває конкретну просторову форму і перетворюється в білок, здатний здійснювати свої гормональні, будівельні, ферментні та інші функції. Для будь-якого організму він є генним продуктом. Саме з нього визначаються різноманітні якості, властивості і прояви генів.

гени
В першу чергу процеси секвенування розроблялися з метою отримання інформації про те, скільки генів має структура молекули ДНК. І, хоча дослідження дозволили вченим далеко просунутися в цьому питанні, дізнатися точну їх кількість поки що не представляється можливим.
Ще кілька років тому передбачалося, що молекули ДНК містять приблизно 100 тис. Генів. Трохи згодом цифра зменшилася до 80 тисяч, а в 1998 р генетиками було заявлено, що в одній ДНК присутня тільки 50 тисяч генів, які є всього лише 3% всієї довжини ДНК. Але вразили останні укладення генетиків. Тепер вони стверджують, що в геном входить 25-40 тисяч згаданих одиниць. Виходить, що за кодування білків відповідає тільки 1,5% хромосомної ДНК.
На цьому дослідження не припинилися. Паралельна команда фахівців генної інженерії встановила, що чисельність генів в одній молекулі становить саме 32 тисячі. Як бачите, отримати остаточну відповідь поки що неможливо. Занадто багато протиріч. Всі дослідники спираються тільки на свої отримані результати.
Чи було еволюціонування?
Незважаючи на те що немає ніяких доказів еволюції молекули (так як будова молекули ДНК крихке і має малий розмір), все ж вченими було висловлено одне припущення. Виходячи з лабораторних даних, вони озвучили версію такого змісту: молекула на початковому етапі своєї появи мала вигляд простого самовоспроизводящегося пептиду, до складу якого входило до 32 амінокислот, що містяться в древніх океанах.
Після самореплікаціі, завдяки силам природного відбору, у молекул з'явилася здатність захищати себе від впливу зовнішніх елементів. Вони стали довше жити і відтворюватися в великих кількостях. Молекули, що знайшли себе в ліпідному міхурі, отримали всі шанси для самовідтворення. В результаті низки послідовних циклів ліпідні бульбашки придбали форму клітинних мембран, а вже далі - всім відомих частинок. Слід зазначити, що сьогодні будь-яку ділянку молекули ДНК являє собою складну і чітко функціонуючу структуру, всі особливості якої вченими до кінця ще не вивчені.

Сучасний світ
Нещодавно вчені з Ізраїлю розробили комп'ютер, якому під силу виконувати трильйони операцій в секунду. Сьогодні це найшвидша машина на Землі. Весь секрет полягає в тому, що інноваційний пристрій функціонує від ДНК. Професори кажуть, що в найближчій перспективі такі комп'ютери зможуть навіть виробляти енергію.
Фахівці з інституту Вейцмана в Реховоті (Ізраїль) рік тому заявили про створення програмованої молекулярної обчислювальної машини, що складається з молекул і ферментів. Ними вони замінили мікрочіпи з кремнію. До теперішнього часу команда ще просунулася вперед. Тепер забезпечити комп'ютер необхідними даними і надати потрібну паливо може всього одна молекула ДНК.
Біохімічні «нанокомп'ютер» - це не вигадка, вони вже існують в природі і проявлені в кожній живій істоті. Але найчастіше вони не справляються людьми. Людина поки що не може оперувати геном якого-небудь рослини, щоб розрахувати, скажімо, число «Пі».
Ідея про використання ДНК для зберігання / обробки даних вперше відвідала світлі голови вчених в 1994 році. Саме тоді для вирішення простий математичної задачі була задіяна молекула. З того моменту ряд дослідницьких груп запропонував різні проекти, що стосуються ДНК-комп'ютерів. Але тут всі спроби ґрунтувалися лише на енергетичній молекулі. Неозброєним оком такий комп'ютер не побачиш, він має вигляд прозорого розчину води, що знаходиться в пробірці. У ньому немає ніяких механічних деталей, а тільки трильйони біомолекулярних пристроїв - і це тільки в одній краплі рідини!
ДНК людини
Який вид у ДНК людини, людям стало відомо в 1953 році, коли вчені вперше змогли продемонструвати світу двухцепочная модель ДНК. За це Кірк і Уотсон отримали Нобелівську премію, так як дане відкриття стало фундаментальним в 20 столітті.
Згодом, звичайно, довели, що не тільки так, як в запропонованому варіанті, може виглядати структурована молекула людини. Провівши більш детальний аналіз ДНК, відкрили А-, В- і левозакрученной форму Z-. Форма А- часто є винятком, так як утворюється тільки в тому випадку, якщо спостерігається недостатність вологи. Але це можливо хіба що при лабораторних дослідженнях, для природного середовища це аномально, в живій клітині такий процес відбуватиметься не може.
Форма В- є класичною і відома як подвійна правозакрученная ланцюг, а ось форма Z- не тільки закручена в зворотному напрямку, вліво, але також має більш зигзагоподібний вигляд. Вченими виділено ще і форма G-квадруплекс. В її структурі не 2, а 4 нитки. На думку генетиків, виникає така форма на тих ділянках, де є надмірна кількість гуаніну.

штучна ДНК
Сьогодні вже існує штучна ДНК, що є ідентичною копією справжньої; вона ідеально повторює структуру природного подвійної спіралі. Але, на відміну від первозданного полинуклеотида, в штучному - всього два додаткових нуклеотиду.
Так як дубляж створювався на основі інформації, отриманої в ході різних досліджень справжньою ДНК, то він також може копіюватися, самовідтворюватися і еволюціонувати. Над створенням такої штучної молекули фахівці працювали близько 20 років. В результаті вийшло дивний винахід, яке може користуватися генетичним кодом так само, як і природна ДНК.
До чотирьох наявних азотистих підстав генетики додали додаткові два, які створили методом хімічної модифікації природних підстав. На відміну від природного, штучна ДНК вийшла досить короткою. Вона містить тільки 81 пару підстав. Проте вона також розмножується і еволюціонує.
Реплікація молекули, отриманої штучним шляхом, Має місце завдяки полімеразної ланцюгової реакції, але поки що це відбувається не самостійно, а через втручання вчених. У згадану ДНК вони самостійно додають необхідні ферменти, поміщаючи її в спеціально підготовлену рідку середу.
Кінцевий результат
На процес і кінцевий результат розвитку ДНК можуть впливати різні чинники, наприклад мутації. Це обумовлює обов'язкове вивчення зразків матерії, щоб результат аналізів був достовірним і надійним. Як приклад можна привести тест на батьківство. Але не може не радувати, що такі казуси, як мутація, зустрічаються рідко. Проте зразки матерії завжди перевіряють, щоб на основі аналізу отримати більш точну інформацію.

ДНК рослин
Завдяки високим технологіям секвенування (HTS) здійснена революція і в області геноміки - виділення ДНК з рослин також можливо. Звичайно, отримання з рослинного матеріалу молекулярної маси ДНК високої якості викликає деякі труднощі, обумовлені великим числом копій мітохондрій і хлоропластів ДНК, а також високим рівнемполісахаридів і фенольних сполук. Для виділення розглянутої нами структури в цьому випадку задіюються найрізноманітніші методи.
Водневий зв'язок в ДНК
За водневу зв'язок в молекулі ДНК відповідає електромагнітне тяжіння, створюване між позитивно зарядженим атомом водню, який приєднаний до електронегативного атома. Дане дипольномувзаємодія не підпадає під критерій хімічного зв'язку. Але вона може здійснитися міжмолекулярної або в різних частинах молекули, т. Е. Внутрішньомолекулярними.

Атом водню приєднується до електронегативного атома, що є донором цих питань. Електронегативним атомом може бути азот, фтор, кисень. Він - шляхом децентралізації - привертає до себе електронну хмару з водневого ядра і робить атом водню зарядженим (частково) позитивно. Так як розмір Н маленький, в порівнянні з іншими молекулами і атомами, заряд виходить також малим.
розшифровка ДНК
Перш ніж розшифрувати молекулу ДНК, вчені спочатку беруть величезна кількість клітин. Для найбільш точної і успішної роботи їх необхідно близько мільйона. Отримані в процесі вивчення результати постійно порівнюють і фіксують. Сьогодні розшифровка генома - це вже не рідкість, а доступна процедура.

Звичайно, розшифровувати геном однієї клітини - це недоцільне заняття. Отримані в ході таких досліджень дані для вчених не уявляють жодного інтересу. Але важливо розуміти, що всі існуючі на даний момент методи декодування, незважаючи на їх складність, недостатньо ефективні. Вони дозволять зчитувати тільки 40-70% ДНК.
Однак гарвардські професори недавно заявили про спосіб, завдяки якому можна розшифрувати 90% генома. Методика заснована на додаванні до виділених клітин молекул-праймерів, за допомогою них і починається реплікація ДНК. Але навіть і цей метод не можна вважати успішним, його ще потрібно доопрацювати, перш ніж відкрито використовувати в науці.
ДНК (дезоксирибонуклеїнова кислота) відноситься до (поряд з РНК), які є полімерами, а точніше - полинуклеотидами (мономер - нуклеотид).
ДНК відповідає за зберігання та передачу при діленні клітин генетичного коду. Саме через молекули ДНК реалізуються спадковість і мінливість. На ДНК синтезуються всі види РНК. Далі різні типи РНК спільно забезпечують синтез білків клітини, т. Е. Реалізують генетичну інформацію.
У клітинах еукаріот переважна кількість ДНК знаходиться в ядрі, де вони утворюють комплекси з особливими білками, в результаті чого утворюються хромосоми. У клітинах прокаріотів існує одна велика кільцева (або лінійна) молекула ДНК (також в комплексі з білками). Крім того в клітинах еукаріот своя ДНК є в мітохондріях і хлоропластах.
У разі ДНК кожен нуклеотид складається з 1) азотистої основи, яке може бути аденін, гуаніном, цитозином або тиміном, 2) дезоксирибози, 3) фосфорної кислоти.
Послідовність нуклеотидів в ланцюжку ДНК визначає первинну структуру молекули. Для ДНК характерна вторинна структура молекули в формі подвійної спіралі (найчастіше правозакрученной). При цьому два ланцюги ДНК з'єднуються між собою водневими зв'язками, утвореними між комплементарними азотистими підставами.
Аденін комплементарен тимін, а гуанін - цитозин. Між аденином і тиміном утворюються дві водневі зв'язку, а між гуаніном і цитозином - три. Таким чином, гуанін і цитозин з'єднані між собою трохи міцніше (хоча водневі зв'язку в принципі слабкі). Кількість зв'язків визначається особливостями будови молекул.
Аденін і гуанін відносяться до пуринів і складаються з двох кілець. Тимін і цитозин відносяться до пірімідіновим підставах, що складається з одного кільця. Таким чином між кістяками (що складаються з чергуються дезоксирибози і фосфорної кислоти) двох ланцюгів ДНК при будь-якій парі нуклеотидів різних ланцюгів завжди існує три кільця (оскільки двухкольцовий пурин завжди комплементарен тільки певного однокольцовому піримідиніл). Це дозволяє зберігати ширину між ланцюгами молекули ДНК однаковою на всьому протязі (приблизно 2,3 нм).
У першому витку спіралі знаходиться приблизно 10 нуклеотидів. Довжина одного нуклеотиду приблизно 0,34 нм. Довжина ж молекул ДНК зазвичай величезна, перевищує мільйони нуклеотидів. Тому, щоб більш компактно розміститися в ядрі клітини, ДНК піддається різного ступеня «сверхспіралізаціі».
При зчитуванні інформації з ДНК (тобто синтезі на ній РНК, цей процес називається транскрипцією) Відбувається деспіралізація (процес зворотний спирализации), два ланцюжки розходяться під дією спеціального ферменту. Водневі зв'язки слабкі, тому поділ і в подальшому зшивання ланцюгів відбувається при малій витраті енергії. РНК синтезується на ДНК згідно все тим же принципом комплементарності. Тільки замість тиміну в РНК аденіну комплементарен урацил.
генетичний код, Записаний на молекулах ДНК, складається з триплетів (послідовностей трьох нуклеотидів), які позначають одну амінокислоту (мономер білка). Однак більша частина ДНК не кодує білок. Значення таких ділянок молекули по-різному, багато в чому до кінця не з'ясовано.
Перед поділом клітини завжди відбувається подвоєння кількості ДНК. Цей процес називається репликацией. Вона носить напівконсервативний характер: ланцюги однієї молекули ДНК розходяться, і на кожній добудовується своя нова комплементарна ланцюг. В результаті з однієї дволанцюгової молекули ДНК виходять дві двухцепочние ДНК, ідентичні першої.
У ДНК полінуклеотидні ланцюга різноспрямовані, т. Е. Там де у одного ланцюга 5 "-кінець (залишок фосфорної кислоти приєднаний до п'ятого атому вуглецю дезоксирибози), в іншої - 3" (вуглець, вільний від фосфорної кислоти).
При реплікації і транскрипції синтез завжди йде в напрямку від 5 "-кінців до 3", так як нові нуклеотиди можуть приєднуватися тільки до вільного 3 "атому вуглецю.
Будова і роль ДНК як речовини, що відповідає за спадкову інформацію, були з'ясовані в 40-50-х роках XX століття. У 1953 році Д. Уотсон і Ф. Крик визначили двухцепочечную структуру ДНК. Раніше Е. Чаргафф з'ясував, що в ДНК кількість тиміну завжди відповідає аденіну, а кількість гуаніну - цитозин.
Схожі статті
-
Немає ніг а ходять 4 літери. Ходять без ніг. Визначення слова годинник в словниках
ЗАГАДКИ Сфінкс Сфінкс задасть вам загадку і в залежності від того, правильно чи ні ви відповісте, благословить або прокляне вас. Як благословення ви можете отримати ресурси, ману, досвід або окуляри пересування. Прокляття може ...
-
Загадка про шкільний дзвінок для дітей
11 Щаслива дитина 16.05.2018 Дорогі читачі, навчання малюків починається ще в дитячому садку. Саме тут закладаються перші основи знань, та й ми завжди поруч, розвиваємо дітей, готуємо їх до школи. А за допомогою загадок ...
-
«Вечір загадок за творами З
Всі ми з дитинства чудово знаємо Самуїла Яковича Маршака - російського радянського поета, який дуже багато книг написав для самих маленьких і допитливих читачів. Саме загадки Маршака залучають дітлахів, і ті з задоволенням ...
-
Битви імперій: Ацтеки Гра ацтеки битви імперій
Куаутемок очолив імперію ацтеків в результаті «ночі печалі». Цей епізод став першим зіткненням правителя з іспанським завойовником Кортесом. «Ніч печалі» з 30 червня на 1 липня 1520 ознаменувалася відступом конкістадорів з ...
-
Ацтеки: битви імперій: керівництва і проходження Ацтеки битви імперій
Вам знайоме слово «марення»? Швидше за все - напевно. Чи може марення бути чудовим? Швидше за все - ні, відповісте ви і ... помилитеся. Повністю забуте творіння російських розробників «Битви імперій: Ацтеки» начисто спростовує ...
-
Різноманітні загадки про вчителя
Загадки про вчителя безумовно сподобаються школярам, адже тих, з ким стикаєшся регулярно, дізнатися найпростіше. Однак ці загадки можна і дати дітям молодшого віку, які вже знайомі з деякими близькими їх сприйняття професіями. Будь-яку ...