DNS-molekula. DNS molekula szerkezete. Örökletes információ: tárolás és átvitel. Genetikai kód. DNS-szál Ahol a DNS-molekulák találhatók
A nukleinsavak mononukleotidokból álló nagy molekulatömegű anyagok, amelyek 3", 5"-os foszfodiészter kötésekkel polimer láncban kapcsolódnak egymáshoz, és bizonyos módon a sejtekben vannak csomagolva.
A nukleinsavak kétféle biopolimerek: ribonukleinsav (RNS) és dezoxiribonukleinsav (DNS). Mindegyik biopolimer nukleotidokból áll, amelyek a szénhidrát-maradékban (ribóz, dezoxiribóz) és az egyik nitrogénbázisban (uracil, timin) különböznek egymástól. E különbségek alapján kapták a nukleinsavak a nevüket.
Dezoxiribonukleinsav szerkezete
A nukleinsavak elsődleges, másodlagos és harmadlagos szerkezettel rendelkeznek.
Elsődleges DNS szerkezet
A DNS elsődleges szerkezetét lineáris polinukleotid láncnak nevezzük, amelyben a mononukleotidok 3", 5"-os foszfodiészter kötésekkel kapcsolódnak egymáshoz. Kiindulási anyag a lánc összeszerelésénél nukleinsav a sejtben egy nukleozid 5"-trifoszfát található, amely a foszforsav β és γ maradékainak eltávolítása eredményeként képes egy másik nukleozid 3" szénatomját hozzákapcsolni. Így az egyik dezoxiribóz 3" szénatomja kovalensen kötődik a másik dezoxiribóz 5" szénatomjához egy foszforsav-maradékon keresztül, és lineáris polinukleotid nukleinsavláncot képez. Innen a név: 3", 5" -foszfodiészter kötések. A nitrogéntartalmú bázisok nem vesznek részt egy lánc nukleotidjainak kombinációjában (1. ábra).
Egy ilyen kapcsolat az egyik nukleotid foszforsavmolekulájának maradéka és egy másik szénhidrátja között a polinukleotid molekula pentóz-foszfát vázának kialakulásához vezet, amelyhez az oldalán egyenként nitrogénbázisok kapcsolódnak. Elrendeződésük sorrendje a nukleinsavmolekulák láncaiban szigorúan specifikus a különböző organizmusok sejtjeire, pl. sajátos jellegű (Chargaff-szabály).
A lineáris DNS-láncnak, amelynek hossza a láncban lévő nukleotidok számától függ, két vége van: az egyik a 3 "vége, és szabad hidroxilcsoportot tartalmaz, a másik, az 5" vége pedig egy foszforsavat tartalmaz. maradékot. A lánc polarizált, iránya 5 "-> 3" és 3 "-> 5" lehet. Kivétel a körkörös DNS.
A DNS genetikai „szövege” kód „szavakból” – kodonoknak nevezett nukleotidhármasokból áll. A DNS azon régióit, amelyek az RNS valamennyi típusának elsődleges szerkezetéről tartalmaznak információt, strukturális géneknek nevezzük.
A polinukleoid DNS-láncok gigantikus méretűek, így bizonyos módon becsomagolódnak a sejtben.
Chargaff (1949) a DNS összetételét tanulmányozva fontos törvényeket állapított meg az egyes DNS-bázisok tartalmára vonatkozóan. Segítettek a DNS másodlagos szerkezetének feltárásában. Ezeket a mintákat Chargaff-szabályoknak nevezzük. Chargaff szabályok
Ezek a szabályok azt jelzik, hogy a DNS felépítésénél nem kell meglehetősen szigorú megfelelést (párosítást) megfigyelni a purin és a pirimidin bázisok között általában, hanem konkrétan a timin és az adenin és a citozin és a guanin között. Ezen szabályok alapján, köztük 1953-ban, Watson és Crick javasolta a DNS másodlagos szerkezetének, a kettős hélixnek nevezett modelljét (ábra). |

A DNS másodlagos szerkezete
A DNS másodlagos szerkezete egy kettős hélix, amelynek modelljét D. Watson és F. Crick javasolta 1953-ban.
A DNS-modell elkészítésének előfeltételei
A kezdeti elemzések azt a benyomást keltették, hogy bármilyen eredetű DNS mind a négy nukleotidot egyenlő moláris mennyiségben tartalmazza. Az 1940-es években azonban E. Chargaff és munkatársai a különböző élőlényekből izolált DNS elemzése eredményeként egyértelműen kimutatták, hogy nitrogénbázisok különböző mennyiségi arányban vannak bennük. Chargaff megállapította, hogy bár ezek az arányok azonosak az azonos típusú organizmus összes sejtjéből származó DNS esetében, a különböző fajokból származó DNS-ek bizonyos nukleotidok tartalmában jelentősen eltérhetnek. Ez arra utalt, hogy a nitrogénbázisok arányának különbségei valamilyen biológiai kóddal hozhatók összefüggésbe. Bár a különböző DNS-mintákban az egyes purin- és pirimidinbázisok aránya eltérőnek bizonyult, az elemzési eredményeket összehasonlítva egy bizonyos minta rajzolódott ki: minden mintában a purinok összmennyisége megegyezett a pirimidinek teljes mennyiségével (A + G = T + C), az adenin mennyisége megegyezett a timin mennyiségével (A = T), és a guanin mennyisége - a citozin mennyiségével (G = C). Az emlőssejtekből izolált DNS általában gazdagabb adeninben és timinben, viszonylag szegényebb guaninban és citozinban, míg a baktériumokban gazdagabb guaninban és citozinban, valamint viszonylag szegényebb adeninben és timinben. Ezek az adatok fontos részét képezték annak a tényanyagnak, amely alapján később felépítették a Watson-Crick DNS szerkezetének modelljét.
A DNS lehetséges szerkezetére vonatkozó másik fontos közvetett jelzés L. Pauling fehérjemolekulák szerkezetére vonatkozó adatai voltak. Pauling kimutatta, hogy egy fehérjemolekulában az aminosavlánc többféle stabil konfigurációja lehetséges. A peptidlánc egyik gyakori konfigurációja, az α-hélix szabályos spirális szerkezet. Ilyen szerkezettel lehetséges a hidrogénkötések kialakulása a lánc szomszédos menetein elhelyezkedő aminosavak között. Pauling 1950-ben leírta a polipeptidlánc α-hélikális konfigurációját, és felvetette, hogy a DNS-molekulák valószínűleg spirális szerkezettel rendelkeznek, amelyet hidrogénkötések rögzítenek.
A DNS-molekula szerkezetéről azonban a legértékesebb információkat a röntgenszerkezeti elemzés eredményei szolgáltatták. A DNS-kristályon áthaladó röntgensugarak diffrakción mennek keresztül, azaz bizonyos irányokba eltérnek. A sugarak eltérülésének mértéke és jellege maguknak a molekuláknak a szerkezetétől függ. A röntgendiffrakciós kép (3. ábra) számos közvetett jelzést ad a tapasztalt szemnek a vizsgált anyag molekuláinak szerkezetére vonatkozóan. A DNS röntgendiffrakciós mintázatának elemzése arra a következtetésre vezetett, hogy a nitrogéntartalmú bázisok (amelyek lapos alakúak) úgy vannak egymásra rakva, mint egy halom lemez. A röntgendiffrakciós minták a kristályos DNS szerkezetének három fő periódusát tárták fel: 0,34, 2 és 3,4 nm.

Watson-Crick DNS-modell
Chargaff analitikai adatai, a Wilkins által nyert röntgendiffrakciós minták és a kémikusok kutatásai alapján, akik információt adtak a molekulában lévő atomok közötti pontos távolságokról, az adott atom kötései közötti szögekről és a molekula méretéről. Watson és Crick a DNS-molekula egyes alkotórészeinek egy bizonyos léptékű fizikai modelljét kezdte építeni, és úgy „illeszteni” őket egymáshoz, hogy a kapott rendszer különböző kísérleti adatoknak feleljen meg. [előadás] .
Már korábban is ismerték, hogy a DNS-lánc szomszédos nukleotidjait foszfodiészter hidak kötik össze, amelyek az egyik nukleotid dezoxiribóz 5'-szénatomját kötik össze a következő nukleotid dezoxiribóz 3'-szénatomjával. Watsonnak és Cricknek nem volt kétsége afelől, hogy a 0,34 nm-es periódus megfelel a DNS-lánc egymást követő nukleotidjai közötti távolságnak. Továbbá feltételezhető, hogy a 2 nm-es periódus megfelel a láncvastagságnak. És annak megmagyarázására, hogy melyik valós szerkezet felel meg a 3,4 nm-es periódusnak, Watson és Crick, akárcsak korábban Pauling, azt javasolta, hogy a lánc spirál formájában csavarodik (vagy pontosabban csavarvonalat alkot, mivel egy spirál ennek szoros értelmében a szót akkor kapjuk, ha a fordulatok a térben nem hengeres, hanem kúpos felületet alkotnak). Ekkor a 3,4 nm-es periódus megfelel a spirál egymást követő fordulatai közötti távolságnak. Egy ilyen spirál lehet nagyon sűrű vagy kissé feszített, azaz a fordulatai lehetnek enyhék vagy meredekek. Mivel a 3,4 nm-es periódus pontosan tízszerese az egymást követő nukleotidok távolságának (0,34 nm), egyértelmű, hogy a hélix minden teljes köre 10 nukleotidot tartalmaz. Ezekből az adatokból Watson és Crick ki tudták számítani egy 2 nm-es hélixre csavart polinukleotid lánc sűrűségét, amelynek menetei közötti távolság 3,4 nm. Kiderült, hogy egy ilyen lánc sűrűsége fele lesz a DNS tényleges sűrűségének, ami már ismert volt. Feltételeznem kellett, hogy a DNS-molekula két szálból áll – ez egy nukleotidok kettős hélixe.
A következő feladat természetesen a kettős csavarvonalat alkotó két áramkör térbeli kapcsolatának tisztázása volt. Miután számos láncelrendezést teszteltek fizikai modelljükön, Watson és Crick úgy találta, hogy minden rendelkezésre álló adat a legjobban illeszkedik egy olyan változathoz, amelyben két polinukleotid hélix ellentétes irányban fut; ebben az esetben cukor- és foszfátmaradékokból álló láncok alkotják a kettős hélix felületét, benne purinok és pirimidinek találhatók. Az egymással szemben elhelyezkedő, két lánchoz tartozó bázisokat páronként hidrogénkötések kötik össze; ezek a hidrogénkötések tartják össze a láncokat, így rögzítve a molekula általános konfigurációját.
A DNS kettős hélixe spirál alakú kötéllétraként képzelhető el, így annak fokai vízszintes helyzetben maradnak. Ekkor két hosszanti kötél cukor- és foszfátmaradék-láncoknak, a keresztrudak pedig hidrogénkötésekkel összekapcsolt nitrogénbázispároknak felelnek meg.
A lehetséges modellek további tanulmányozása eredményeként Watson és Crick arra a következtetésre jutott, hogy minden "rudat" egy purinból és egy pirimidinből kell állnia; 2 nm periódussal (ami a kettős hélix átmérőjének felel meg) nem lenne elég hely a két purinnak, és a két pirimidin sem lenne elég közel egymáshoz ahhoz, hogy megfelelő hidrogénkötéseket hozzon létre. Egy részletes modell alapos tanulmányozása kimutatta, hogy az adenin és a citozin, amelyek megfelelő méretű kombinációt alkotnak, még mindig nem helyezhetők el úgy, hogy közöttük hidrogénkötés alakuljon ki. Hasonló jelentések kényszerítették a guanin-timin kombináció kizárását, míg az adenin-timin és guanin-citozin kombinációk meglehetősen elfogadhatóak voltak. A hidrogénkötések természete olyan, hogy az adenin a timinnel, a guanin pedig a citozinnal alkot párt. Ez a specifikus bázispárosítás koncepciója lehetővé tette a „Chargaff-szabály” magyarázatát, amely szerint bármely DNS-molekulában az adenin mennyisége mindig megegyezik a timin-tartalommal, a guanin mennyisége pedig a citozin mennyiségével. Két hidrogénkötés jön létre az adenin és a timin között, három pedig a guanin és a citozin között. Az egyik láncban az egyes adeninekkel szembeni hidrogénkötések képződésének e sajátossága miatt a timin a másik láncban található; hasonlóképpen minden guanin ellen csak citozin található. Így a láncok komplementerek egymással, vagyis az egyik lánc nukleotidsorrendje egyértelműen meghatározza a szekvenciát a másikban. A két lánc ellentétes irányban fut, és terminális foszfátcsoportjaik a kettős hélix ellentétes végén találhatók.
Kutatásaik eredményeként 1953-ban Watson és Crick javasolta a DNS-molekula szerkezetének modelljét (3. ábra), amely a mai napig is releváns. A modell szerint egy DNS-molekula két komplementer polinukleotid láncból áll. Mindegyik DNS-szál több tízezer nukleotidból álló polinukleotid. Ebben a szomszédos nukleotidok szabályos pentóz-foszfát vázat alkotnak a foszforsav-maradék és a dezoxiribóz erős kovalens kötéssel történő összekapcsolódása miatt. Ebben az esetben az egyik polinukleotid lánc nitrogéntartalmú bázisai szigorúan meghatározott sorrendben helyezkednek el a másik lánc nitrogéntartalmú bázisaival szemben. A nitrogéntartalmú bázisok váltakozása a polinukleotid láncban szabálytalan.
A nitrogénbázisok elhelyezkedése a DNS-láncban komplementer (a görög "komplement" szóból - addíció), azaz. az adenin (A) ellen mindig van timin (T), a guanin ellen (G) pedig csak a citozin (C). Ez annak köszönhető, hogy A és T, valamint G és C szigorúan megfelelnek egymásnak, i.e. kiegészítik egymást. Ilyen levelezést adnak kémiai szerkezete bázisok, lehetővé téve hidrogénkötések kialakulását egy purin és pirimidin párban. Két kapcsolat van A és T között, három pedig G és C között. Ezek a kötések biztosítják a DNS-molekula részleges stabilizálását a térben. Ebben az esetben a kettős hélix stabilitása egyenesen arányos a G≡C kötések számával, amelyek stabilabbak az A = T kötésekhez képest.
Az egyik DNS-szálban a nukleotidok elrendeződésének ismert szekvenciája lehetővé teszi a komplementaritás elve szerint a másik szál nukleotidjainak megállapítását.
Ezenkívül azt találták, hogy aromás szerkezetű nitrogéntartalmú bázisok vizesoldat egymás felett helyezkednek el, mintegy érmehalmazt alkotva. A szerves molekulák halmozásának ezt a folyamatát halmozásnak nevezik. A vizsgált Watson-Crick modell DNS molekulájának polinukleotid láncai hasonló fizikai-kémiai állapotúak, nitrogéntartalmú bázisaik érmeköteg formájában helyezkednek el, amelyek síkjai között van der Waals kölcsönhatások (halmozási kölcsönhatások) keletkeznek.
Hidrogénkötések A komplementer bázisok között (vízszintesen) és a polinukleotid láncban a bázisok síkjai között a van der Waals erők hatására (függőlegesen) egymásra halmozódó kölcsönhatás további stabilizációt biztosít a DNS-molekulának a térben.
Mindkét lánc cukor-foszfát gerince kifelé, az alapjai befelé, egymás felé néznek. A DNS-ben a láncok iránya antiparallel (az egyik iránya 5 "-> 3", a másiké - 3 "-> 5", azaz az egyik lánc 3 "vége a másik 5" végével szemben van .). A láncok jobb oldali spirálokat alkotnak, amelyeknek közös tengelye van. A hélix egy fordulata 10 nukleotidból áll, a tekercs mérete 3,4 nm, az egyes nukleotidok magassága 0,34 nm, a hélix átmérője 2,0 nm. Az egyik szálnak a másik körül forgásának eredményeként a DNS kettős hélix egy nagy (kb. 20 Å átmérőjű) és egy kis barázda (kb. 12 Å) képződik. A Watson-Crick kettős hélixnek ezt a formáját később B-formának nevezték. A sejtekben a DNS általában B-formában létezik, ami a legstabilabb forma.
DNS funkciók
A javasolt modell sok mindent megmagyaráz biológiai tulajdonságait dezoxiribonukleinsav, beleértve a genetikai információ tárolását és a gének sokféleségét, amelyet 4 nukleotid szekvenciális kombinációinak széles választéka biztosít, valamint a genetikai kód létezésének ténye, az önreprodukciós képesség és a genetikai információ átvitele, feltéve, hogy a replikációs folyamat, valamint a genetikai információ megvalósítása fehérjék és minden más enzimfehérjék által képzett vegyület formájában.
A DNS alapvető funkciói.
- A DNS a genetikai információ hordozója, amit a genetikai kód meglétének ténye biztosít.
- A genetikai információ szaporodása és átvitele sejtek és organizmusok generációiban. Ezt a funkciót a replikációs folyamat biztosítja.
- Genetikai információ realizálása fehérjék formájában, valamint bármely más, enzimfehérjék segítségével képződő vegyület formájában. Ezt a funkciót az átírási és fordítási folyamatok biztosítják.

A kettős szálú DNS szerveződési formái
A DNS többféle kettős hélixet képezhet (4. ábra). Jelenleg már hat forma ismert (A-tól E-ig és Z-forma).
A DNS szerkezeti formái, amint azt Rosalind Franklin megállapította, a nukleinsavmolekula vízzel való telítettségétől függenek. A DNS-szálak röntgensugaras szerkezeti analízissel végzett vizsgálatai során kimutatták, hogy a röntgendiffrakciós mintázat radikálisan függ attól, hogy ennek a rostnak milyen relatív páratartalom mellett, milyen víztelítettségi fokon történik a kísérlet. Ha a szál megfelelően telített volt vízzel, akkor egy röntgenfelvételt készítettek. Szárításkor teljesen más röntgenkép jelent meg, ami nagyon eltért a nagy nedvességtartalmú rost röntgenképétől.
A magas páratartalmú DNS-molekulát B-formának nevezik... Fiziológiás körülmények között (alacsony sókoncentráció, magas hidratáltság) a DNS domináns szerkezeti típusa a B-forma (a kettős szálú DNS fő formája a Watson-Crick modell). Egy ilyen molekula hélix-emelkedése 3,4 nm. Körönként 10 komplementer pár van csavart "érmék" - nitrogéntartalmú bázisok - formájában. A kötegeket hidrogénkötések tartják a verem két egymással szemben lévő "érméje" között, és a foszfodiészter gerinc két szalagjába vannak "tekerve", amelyek jobb oldali spirálba vannak csavarva. A nitrogéntartalmú bázisok síkjai merőlegesek a spirál tengelyére. A szomszédos komplementer párokat egymáshoz képest 36°-kal elforgatjuk. A hélix átmérője 20 Å, a purin nukleotid 12 Å-t, a pirimidin nukleotid pedig 8 Å-t foglal el.
Az alacsonyabb páratartalmú DNS-molekulát A-formának nevezik... Az A-forma kevésbé magas hidratáltság és nagyobb Na + vagy K + ion tartalom mellett jön létre. Ez a szélesebb jobbkezes konformáció körönként 11 bázispárt tartalmaz. A nitrogéntartalmú alapok síkjai erősebben hajlanak a spiráltengelyhez, a normáltól a spiráltengelyhez képest 20°-kal eltérnek. Ebből következik, hogy egy 5 Å átmérőjű belső üreg van jelen. A szomszédos nukleotidok távolsága 0,23 nm, a tekercs hossza 2,5 nm, a hélix átmérője 2,3 nm.
Eredetileg a DNS A-formáját kevésbé tartották fontosnak. Később azonban világossá vált, hogy a DNS A-formájának, valamint a B-formájának óriási biológiai jelentősége van. A templát-primer komplexben lévő RNS-DNS hélixnek A-formája, valamint RNS-RNS-hélixe és RNS hajtűszerkezetei vannak (a ribóz 2'-hidroxilcsoportja nem teszi lehetővé, hogy az RNS-molekulák B-formát alkossanak) . A DNS A-formáját vita tárgyaként találták meg. Azt találták, hogy a DNS A-formája 10-szer jobban ellenáll az UV-sugárzásnak, mint a B-forma.
Az A-formát és a B-formát a DNS kanonikus formáinak nevezzük.
C-E formák jobbkezesek is, kialakulásuk csak speciális kísérletekben figyelhető meg, és úgy tűnik, in vivo nem léteznek. A C-formájú DNS szerkezete hasonló a B-DNS-hez. A bázispárok száma körönként 9,33, a hélix hossza 3,1 nm. Az alappárok a tengelyre merőleges helyzethez képest 8 fokos szögben vannak megdöntve. A barázdák mérete hasonló a B-DNS barázdáihoz. Ebben az esetben a fő horony valamivel sekélyebb, a mellékhorony pedig mélyebb. A természetes és szintetikus DNS-polinukleotidok átjuthatnak a C-formába.
| 1. táblázat: Egyes DNS-szerkezetek jellemzői | |||
| Spirális típus | A | B | Z |
| Spirális lépés | 0,32 nm | 3,38 nm | 4,46 nm |
| Spirális csavarás | Jobb | Jobb | Bal |
| Alappárok körönként | 11 | 10 | 12 |
| Alapsíkok közötti távolság | 0,256 nm | 0,338 nm | 0,371 nm |
| Glikozid kötés konformációja | anti | anti | anti-C syn-g |
| Furanóz gyűrű felépítése | C3 "-endo | C2 "-endo | C3 "-endo-G C2 "-endo-c |
| Horonyszélesség kicsi/nagy | 1,11 / 0,22 nm | 0,57 / 1,17 nm | 0,2/0,88 nm |
| Horonymélység, kicsi/nagy | 0,26 / 1,30 nm | 0,82 / 0,85 nm | 1,38 / 0,37 nm |
| Spirál átmérő | 2,3 nm | 2,0 nm | 1,8 nm |
A DNS szerkezeti elemei
(nem kanonikus DNS-struktúrák)
A DNS szerkezeti elemei közé tartoznak a szokatlan struktúrák, amelyeket néhány speciális szekvencia korlátoz:
|

Z alakú DNS 1979-ben fedezték fel a hexanukleotid d (CG) 3 - tanulmányozása során. Alexander Rich MIT professzor és kollégái fedezték fel. A Z-forma a DNS egyik legfontosabb szerkezeti elemévé vált annak köszönhetően, hogy képződését olyan DNS-régiókban figyelték meg, ahol a purinok pirimidinekkel váltakoznak (például 5'-HCGCH-3'), vagy az 5'-es ismétlődésekben. -CHCH-3' metilezett citozint tartalmaz. A Z-DNS kialakulásának és stabilizálásának elengedhetetlen feltétele volt a purin nukleotidok jelenléte a szin-konformációban, felváltva a pirimidin bázisokkal az antikonformációban.
A természetes DNS-molekulák főleg a megfelelő B-formában léteznek, ha nem tartalmaznak (CH) n típusú szekvenciákat. Ha azonban ilyen szekvenciák szerepelnek a DNS-ben, akkor ezek a régiók, amikor az oldat ionerőssége megváltozik, vagy a foszfodiészterváz negatív töltését semlegesítő kationok Z-formává alakulhatnak át, míg a lánc többi DNS-régiója megmarad. a klasszikus B-formában. Az ilyen átmenet lehetősége azt jelzi, hogy a DNS kettős hélixben lévő két lánc dinamikus állapotban van, és egymáshoz képest letekercselhet, átmegy a jobb formából a bal oldaliba és fordítva. Az ilyen labilitás biológiai következményei, amelyek lehetővé teszik a DNS szerkezetének konformációs átalakulását, még nem teljesen ismertek. Úgy gondolják, hogy a Z-DNS régiók szerepet játszanak egyes gének expressziójának szabályozásában, és részt vesznek a genetikai rekombinációban.
A DNS Z-formája egy bal oldali kettős hélix, amelyben a foszfodiészter gerince cikkcakkosan helyezkedik el a molekula tengelye mentén. Innen a molekula neve (cikkcakk) -DNS. A Z-DNS a legkevésbé csavarodott (fordulónként 12 bázispár) és a legvékonyabb a természetben. A szomszédos nukleotidok távolsága 0,38 nm, a tekercs hossza 4,56 nm, a Z-DNS átmérője 1,8 nm. Ezenkívül ennek a DNS-molekulának a megjelenése egyetlen barázda jelenléte is megkülönböztethető.
A DNS Z-formáját prokarióta és eukarióta sejtekben találták meg. Jelenleg olyan antitesteket szereztek, amelyek meg tudják különböztetni a DNS Z-formáját a B-formától. Ezek az antitestek a Drosophila (Dr. melanogaster) nyálmirigysejtjeinek óriáskromoszómáinak meghatározott régióihoz kötődnek. A kötődési reakció könnyen követhető e kromoszómák szokatlan szerkezete miatt, amelyben a sűrűbb régiók (korongok) ellentétben állnak a kevésbé sűrű régiókkal (interkorongokkal). A Z-DNS szakaszai a sávközi sávokban helyezkednek el. Ebből az következik, hogy a Z-alak természetes körülmények között valóban létezik, bár a Z-forma egyes szakaszainak méretei még nem ismertek.

(shifters) a DNS legismertebb és leggyakrabban előforduló bázisszekvenciái. A palindrom egy olyan szó vagy kifejezés, amely ugyanúgy balról jobbra és fordítva olvasható. Példák az ilyen szavakra vagy kifejezésekre: SHALASH, KAZAK, POTOP, AND ROSE FALLED AZOR'S Paw. A DNS-régiókra alkalmazva ez a kifejezés (palindrom) a nukleotidok ugyanazt a váltakozását jelenti a lánc mentén jobbról balra és balról jobbra (mint a "kunyhó" szó betűi stb.).
A palindromot két DNS-szálhoz képest másodrendű szimmetriával rendelkező bázisszekvenciák fordított ismétlődéseinek jelenléte jellemzi. Az ilyen szekvenciák teljesen érthető okból önmagukat kiegészítik, és hajlamosak hajtű- vagy kereszt alakú struktúrákat kialakítani (ábra). A hajtűk segítenek a szabályozó fehérjéknek felismerni azt a helyet, ahol a kromoszóma DNS genetikai szövege le van írva.
Azokban az esetekben, amikor egy fordított ismétlődés ugyanazon a DNS-szálon van jelen, ezt a szekvenciát tükörismétlődésnek nevezzük. A tükör ismétlődések nem rendelkeznek az önkiegészítő tulajdonságokkal, ezért nem képesek hajtű vagy kereszt alakú struktúrák kialakítására. Az ilyen típusú szekvenciák gyakorlatilag minden nagy DNS-molekulában megtalálhatók, és néhány bázispártól több ezer bázispárig terjedhetnek.
A palindromok keresztes struktúrák formájában való jelenléte eukarióta sejtekben nem bizonyított, bár számos kereszt alakú struktúrát találtak in vivo E. coli sejtekben. Az önkomplementer szekvenciák jelenléte az RNS-ben vagy az egyszálú DNS-ben a fő oka annak, hogy a nukleinlánc az oldatokban egy bizonyos térbeli struktúrába hajtódik, amelyet sok "hajtű" képződése jellemez.

H-formájú DNS egy hélix, amelyet három DNS-szál alkot - a DNS-hármas hélix. Ez a Watson-Crick kettős hélix komplexe a harmadik egyszálú DNS-száljal, amely beleillik annak nagy barázdájába, az úgynevezett Hoogsteen pár kialakulásával.
Az ilyen triplex kialakulása a DNS kettős hélix felhajtása eredményeként következik be oly módon, hogy szakaszának fele kettős hélix formájában marad, a másik fele pedig elválik. Ebben az esetben az egyik leválasztott spirál új szerkezetet alkot a kettős spirál első felével - egy hármas spirállal, a második pedig strukturálatlannak bizonyul, egyszálú szakasz formájában. Ennek a szerkezeti átmenetnek a sajátossága az éles függés a közeg pH-jától, amelynek protonjai stabilizálják az új szerkezetet. Ennek a tulajdonságnak köszönhetően új szerkezet kapta a DNS H-formájának a nevét, amelynek kialakulását homopurin-homopirimidin régiókat tartalmazó szuperspirálos plazmidokban találták meg, amelyek tükörismétlődésnek számítanak.
További vizsgálatok során néhány homopurin-homopirimidin kettős szálú polinukleotid szerkezeti átmenetének lehetősége egy háromszálú szerkezet kialakításával, amely tartalmazza:
- egy homopurin és két homopirimidin szál ( Py-Pu-Py triplex) [Hoogsteen interakció].
A Py-Pu-Py triplex alkotó blokkjai a CGC + és a TAT izomorf kanonikus triádjai. A triplex stabilizálásához a CGC + triád protonálódása szükséges, ezért ezek a triplexek az oldat pH-jától függenek.
- egy homopirimidin és két homopurin szál ( Py-Pu-Pu triplex) [fordított Hoogsteen interakció].
A Py-Pu-Pu triplex alkotó blokkjai kanonikus izomorfok a CGG és a TAA triáddal. A Py-Pu-Pu triplexek lényeges tulajdonsága, hogy stabilitásuk kettős töltésű ionok jelenlététől függ, és a különböző szekvenciájú triplexek stabilizálásához különböző ionokra van szükség. Mivel a Py-Pu-Pu triplexek képződése nem igényli az őket alkotó nukleotidok protonálását, az ilyen triplexek semleges pH-n létezhetnek.
Megjegyzés: a közvetlen és fordított Hoogsteen kölcsönhatást az 1-metiltimin szimmetriája magyarázza: a 180°-os elforgatás azt a tényt eredményezi, hogy az O4 atomot az O2 atom helyettesíti, miközben a hidrogénkötések rendszere megmarad.
Kétféle hármas spirál létezik:
- párhuzamos hármas hélixek, amelyekben a harmadik szál polaritása egybeesik a Watson-Crick duplex homopurin láncának polaritásával
- antiparallel tripla hélixek, amelyekben a harmadik és a homopurin lánc polaritása ellentétes.

G-quadruplex- 4-hélix DNS. Ilyen szerkezet akkor jön létre, ha négy guanin van, amelyek alkotják az úgynevezett G-quadruplexet - négy guanin körtáncát.
Az első utalások az ilyen struktúrák kialakulásának lehetőségére jóval Watson és Crick áttörő munkája előtt érkeztek - még 1910-ben. Aztán Ivar Bang német vegyész felfedezte, hogy a DNS egyik összetevője - a guanózsav - magas koncentrációban géleket képez, míg a DNS többi összetevője nem rendelkezik ezzel a tulajdonsággal.
1962-ben röntgendiffrakciós módszerrel sikerült megállapítani ennek a gélnek a sejtszerkezetét. Kiderült, hogy négy guaninmaradékból áll, amelyek körbe kapcsolják egymást, és jellegzetes négyzetet alkotnak. Középen a kötést fémion (Na, K, Mg) tartja. Ugyanezek a struktúrák kialakulhatnak a DNS-ben is, ha sok guanint tartalmaz. Ezek a lapos négyzetek (G-kvartettek) egymásra vannak rakva, hogy meglehetősen stabil, sűrű struktúrákat (G-quadruplex) képezzenek.
Négy különálló DNS-szál négyszálú komplexekké fonódhat össze, de ez inkább kivétel. Gyakrabban előfordul, hogy egyetlen nukleinsavszálat egyszerűen csomóba kötnek, jellegzetes megvastagodásokat hozva létre (például a kromoszómák végein), vagy a kettős szálú DNS helyi kvadrupplexet alkot valamilyen guaninban gazdag régióban.
A legtöbbet tanulmányozott kvadrupplexek létezése a kromoszómák végén - a telomereken és az onkopromotorokban. Mindeddig azonban nem ismert az ilyen DNS lokalizációjának teljes megértése az emberi kromoszómákban.
Mindezek a szokatlan DNS-struktúrák lineáris formában instabilok a DNS B-formájához képest. A DNS azonban gyakran topológiai stressz körkörös formájában létezik, amikor az úgynevezett szupertekercselés. Ilyen körülmények között könnyen kialakulnak nem kanonikus DNS-struktúrák: Z-formák, keresztek és hajtűk, H-formák, guanin-kvadrupplexek és i-motívum.
- Szupertekercses forma - akkor figyelhető meg, amikor a pentóz-foszfát gerinc károsítása nélkül szabadul fel a sejtmagból. Szupercsavart zárt gyűrű alakú. Szupertekercses állapotban a DNS kettős hélix legalább egyszer „magára csavarodik”, azaz legalább egy szupertekercset tartalmaz (nyolcas alakot vesz fel).
- A DNS ellazult állapota egyetlen megszakítással (egy szál törésével) figyelhető meg. Ebben az esetben a szupertekercsek eltűnnek, és a DNS zárt gyűrűt ölt.
- A DNS lineáris formája - akkor figyelhető meg, amikor a kettős hélix két szála eltörik.
DNS harmadlagos szerkezet
DNS harmadlagos szerkezet egy kétszálú molekula térbeli további csavarodása - szupertekervénye - eredményeként jön létre. A DNS-molekula szuperspirálozása az eukarióta sejtekben, a prokariótáktól eltérően, fehérjékkel alkotott komplexek formájában történik.
Szinte az összes eukarióta DNS megtalálható a sejtmagok kromoszómáiban, ennek csak kis része található a mitokondriumokban, valamint a növényekben és a plasztidokban. Az eukarióta sejtek kromoszómáinak (beleértve az emberi kromoszómákat is) fő anyaga a kromatin, amely kettős szálú DNS-ből, hisztonból és nem hiszton fehérjékből áll.
Kromatin hiszton fehérjék
A hisztonok egyszerű fehérjék, amelyek a kromatin 50%-át teszik ki. Az összes vizsgált állati és növényi sejtben a hisztonok öt fő osztályát találták: H1, H2A, H2B, H3, H4, méretükben eltérőek, aminosav összetételés a töltés mértéke (mindig pozitív).
Az emlős hiszton H1 egyetlen polipeptidláncból áll, amely körülbelül 215 aminosavból áll; más hisztonok mérete 100 és 135 aminosav között változik. Mindegyik spirálozott és körülbelül 2,5 nm átmérőjű gömbölyűvé van csavarva, szokatlanul nagy mennyiségű pozitív töltésű aminosavat tartalmaznak, lizint és arginint. A hisztonok lehetnek acetilezve, metilezve, foszforilálva, poli(ADP)-ribozilálva, a H2A és H2B hisztonok pedig kovalensen kapcsolódnak az ubiquitinhez. Az ilyen módosítások szerepe a hisztonok szerkezetének kialakításában és funkcióinak kialakításában még nem teljesen tisztázott. Feltételezhető, hogy ez az a képességük, hogy kölcsönhatásba lépnek a DNS-sel, és biztosítják a gének működésének szabályozásának egyik mechanizmusát.
A hisztonok elsősorban a DNS negatív töltésű foszfátcsoportjai és a hisztonok pozitív töltésű lizin- és argininmaradékai között képződő ionos kötéseken (sóhidak) keresztül lépnek kölcsönhatásba a DNS-sel.
Nem hiszton kromatin fehérjék
A hisztonokkal ellentétben a nem hiszton fehérjék nagyon változatosak. A DNS-kötő nem hiszton fehérjék 590 különböző frakcióját izolálták. Savas fehérjéknek is nevezik őket, mivel szerkezetükben a savas aminosavak dominálnak (polianionok). A kromatin aktivitás specifikus szabályozása számos nem hiszton fehérjéhez kapcsolódik. Például a DNS replikációjához és expressziójához szükséges enzimek átmenetileg kötődhetnek a kromatinhoz. Más fehérjék például, amelyek különböző szabályozási folyamatokban vesznek részt, csak meghatározott szövetekben vagy a differenciálódás bizonyos szakaszaiban kötődnek a DNS-hez. Mindegyik fehérje komplementer egy meghatározott DNS-nukleotidszekvenciával (DNS-hely). Ez a csoport a következőket tartalmazza:
- helyspecifikus cink ujjfehérjék családja. Minden cinkujj felismer egy specifikus helyet, amely 5 nukleotidpárból áll.
- helyspecifikus fehérjék családja - homodimerek. Egy ilyen fehérje DNS-sel érintkező fragmense hélix-turn-helix szerkezetű.
- a nagy mobilitású gélfehérjék (HMG proteinek) olyan szerkezeti és szabályozó fehérjék csoportját alkotják, amelyek folyamatosan kapcsolatban állnak a kromatinnal. Molekulatömege kisebb, mint 30 kDa, és magas töltött aminosav-tartalom jellemzi őket. Alacsony molekulatömegük miatt a HMG fehérjék rendkívül mobilak a poliakrilamid gélelektroforézis során.
- replikációs, transzkripciós és javítási enzimek.
A DNS és az RNS szintézisében részt vevő szerkezeti, szabályozó fehérjék és enzimek részvételével a nukleoszómaszál erősen kondenzált fehérjék és DNS komplexekké alakul. A kapott szerkezet 10 000-szer rövidebb, mint az eredeti DNS-molekula.

Kromatin
A kromatin fehérjék komplexe nukleáris DNS-sel és szervetlen anyagokkal. A kromatin nagy része inaktív. Szorosan csomagolt, kondenzált DNS-t tartalmaz. Ez heterokromatin. Különbséget kell tenni a konstitutív, genetikailag inaktív kromatin (műhold DNS), amely nem expresszálódott régiókból áll, és a fakultatív - több generáción át inaktív, de bizonyos körülmények között expresszálódni képes.
Az aktív kromatin (euchromatin) nem kondenzált, azaz. kevésbé szorosan csomagolva. Különböző cellákban a tartalma 2 és 11% között mozog. Az agy sejtjeiben ez a legtöbb - 10-11%, a máj sejtjeiben - 3-4 és a vesékben - 2-3%. Az euchromatin aktív transzkripciója figyelhető meg. Sőt, szerkezeti felépítése lehetővé teszi a DNS-ben rejlő azonos genetikai információ felhasználását ez a fajta szervezetben, eltérően a speciális sejtekben.
Elektronmikroszkópban a kromatin képe egy gyöngyhöz hasonlít: körülbelül 10 nm méretű gömbszerű vastagodások, amelyeket fonalszerű hidak választanak el. Ezeket a gömb alakú megvastagodásokat nukleoszómáknak nevezzük. A nukleoszóma a kromatin szerkezeti egysége. Mindegyik nukleoszóma egy 146 bázispár hosszúságú, szupertekercses DNS-szegmenst tartalmaz, amely a nukleoszómamagban 1,75 bal oldali fordulattal van feltekerve. A nukleoszómális mag egy hisztonoktamer, amely H2A, H2B, H3 és H4 hisztonokból áll, mindkét típusú két molekulából (9. ábra), amely 11 nm átmérőjű és 5,7 nm vastag korongnak tűnik. Az ötödik hiszton, a H1, nem része a nukleoszómális magnak, és nem vesz részt a hisztonoktamer DNS-tekercselési folyamatában. A DNS-sel érintkezik, ahol a kettős hélix belép és kilép a nukleoszóma magjából. Ezek interkortikális (linker) DNS-régiók, amelyek hossza a sejttípustól függően 40-50 nukleotidpár között változik. Ennek eredményeként a nukleoszómában lévő DNS-fragmens hossza is változik (186-196 nukleotidpár).
A nukleoszóma a DNS körülbelül 90%-át tartalmazza, a többi része a linker. Úgy gondolják, hogy a nukleoszómák a "néma" kromatin töredékei, és a linker aktív. A nukleoszómák azonban kibontakozhatnak és lineárissá válhatnak. A kibontott nukleoszómák már aktív kromatin. Így nyilvánul meg egyértelműen a funkció függése a szerkezettől. Feltételezhető, hogy minél több kromatin van a globuláris nukleoszómák összetételében, annál kevésbé aktív. Nyilvánvaló, hogy a különböző sejtekben a nyugvó kromatin egyenlőtlen aránya összefügg az ilyen nukleoszómák számával.
Az elektronmikroszkópos fényképeken az izolálási körülményektől és a nyújtás mértékétől függően a kromatin nemcsak egy hosszú fonalnak tűnhet megvastagodásokkal - nukleoszóma "gyöngyökkel", hanem rövidebb és sűrűbb, 30 átmérőjű fibrillának (szálnak) is. nm, amelynek képződése a DNS linker régiójához kötődő H1 hiszton és a H3 hiszton kölcsönhatás során figyelhető meg, ami menetenként hat nukleoszómából álló hélix további csavarodásához vezet egy 30 nm átmérőjű szolenoid kialakulásával. Ebben az esetben a hisztonfehérje számos gén transzkripcióját zavarhatja, és így szabályozhatja azok aktivitását.
A DNS és a hisztonok fent leírt kölcsönhatásainak eredményeként a DNS kettős hélixének egy 186 bázispárból álló, átlagosan 2 nm átmérőjű és 57 nm hosszúságú szegmense átalakul 10 nm átmérőjű hélixmé és egy hossza 5 nm. Ennek a spirálnak egy 30 nm átmérőjű szálra történő ezt követő összenyomásával a kondenzáció mértéke hatszorosára nő.
Végül a DNS duplex öt hisztonnal való feltöltése a DNS 50-szeres kondenzációját eredményezi. Azonban még ilyen nagyfokú kondenzáció sem magyarázhatja meg a metafázis kromoszómában a DNS csaknem 50-100 ezerszeres sűrűségét. Sajnos a további kromatinpakolás részletei a metafázisos kromoszómáig még nem ismertek, ezért ennek a folyamatnak csak az általános jellemzői jöhetnek számításba.
DNS tömörítési szint a kromoszómákban
Minden DNS-molekula külön kromoszómába van csomagolva. Az emberi diploid sejtek 46 kromoszómát tartalmaznak, amelyek a sejtmagban találhatók. Egy sejt összes kromoszómájának DNS-ének teljes hossza 1,74 m, de a mag átmérője, amelyben a kromoszómák vannak, milliószor kisebb. A kromoszómákban és a sejtmag kromoszómáiban található DNS ilyen kompakt tömörülését számos hiszton és nem hiszton fehérje biztosítja, amelyek egy meghatározott szekvenciában lépnek kölcsönhatásba a DNS-sel (lásd fent). A kromoszómákban lévő DNS tömörítése lehetővé teszi a lineáris méretének körülbelül 10 000-szeres csökkentését - hagyományosan 5 cm-ről 5 mikronra. A tömörítésnek több fokozata van (10. ábra).


- A DNS kettős hélix egy negatív töltésű molekula, amelynek átmérője 2 nm és hossza néhány cm.
- nukleoszómális szint- a kromatin elektronmikroszkópban "gyöngyök" - nukleoszómák - "egy húron" álló láncának néz ki. A nukleoszóma egy univerzális szerkezeti egység, amely mind az euchromatinban, mind a heterokromatinban, az interfázisú magban és a metafázisú kromoszómákban megtalálható.
A tömörítés nukleoszomális szintjét speciális fehérjék - hisztonok - biztosítják. Nyolc pozitív töltésű hisztondomén alkotja a nukleoszóma magját (magját), amely köré a negatív töltésű DNS-molekula feltekercselődik. Ez hétszeres rövidülést eredményez, miközben az átmérő 2-ről 11 nm-re nő.
- mágnesszelep szint
A kromoszómaszerveződés szolenoid szintjét a nukleoszóma filamentum elcsavarodása jellemzi, és ebből vastagabb, 20-35 nm átmérőjű fibrillák - szolenoidok vagy szuperbidek - képződnek. A mágnesszelep osztásköze 11 nm, fordulatonként körülbelül 6-10 nukleoszóma van. A szolenoidos pakolást valószínűbbnek tartják, mint a szuperbidet, amely szerint egy 20-35 nm átmérőjű kromatinszál egy szemcsékből vagy szuperbidekből álló lánc, amely egyenként nyolc nukleoszómából áll. Solenoid szinten a DNS lineáris mérete 6-10-szeresére csökken, az átmérő 30 nm-re nő.
- hurokszint
A hurokszintet nem hiszton helyspecifikus DNS-kötő fehérjék biztosítják, amelyek felismernek és kötődnek specifikus DNS-szekvenciákhoz, körülbelül 30-300 kb méretű hurkokat képezve. A hurok biztosítja a génexpressziót, azaz az E. a hurok nemcsak szerkezeti, hanem funkcionális képződmény is. A rövidülés ezen a szinten 20-30 alkalommal történik. Az átmérő 300 nm-re nő. A kétéltű petesejtekben "lámpakefe" típusú hurokszerű struktúrák láthatók a citológiai készítményeken. Ezek a hurkok láthatóan szuperspirálosak, és olyan DNS-doméneket képviselnek, amelyek valószínűleg megfelelnek a kromatin transzkripció és replikáció egységeinek. Specifikus fehérjék rögzítik a hurkok alapjait és esetleg egyes belső régióikat. A hurok alakú domén szerveződés elősegíti a metafázisú kromoszómák kromatinjának magasabb rendű spirális struktúrákká való feltekeredését.
- domain szinten
A kromoszóma szerveződésének doménszintjét nem vizsgálták kellőképpen. Ezen a szinten a hurokdomének kialakulása figyelhető meg - a 25-30 nm vastagságú szálak (fibrillák) szerkezetei, amelyek 60% fehérjét, 35% DNS-t és 5% RNS-t tartalmaznak, gyakorlatilag láthatatlanok a sejt minden fázisában a mitózis kivételével, és némileg véletlenszerűen oszlanak el a sejtmagban. A kétéltű petesejtekben "lámpakefe" típusú hurokszerű struktúrák láthatók a citológiai készítményeken.
A hurokdomének bázisukkal az intranukleáris fehérjemátrixhoz kapcsolódnak az úgynevezett beágyazott kapcsolódási helyeken, amelyeket gyakran MAR / SAR szekvenciáknak neveznek (MAR, az angol mátrixhoz kapcsolódó régióból; SAR, az angol scaffold kapcsolódási régiókból) - DNS több száz bázispár hosszúságú töredékek, amelyekre jellemző a magas (> 65%) A/T bázispár tartalom. Úgy tűnik, hogy minden tartomány egyetlen replikációs origóval rendelkezik, és önálló szupertekercses egységként működik. Bármely hurkolt tartomány sok transzkripciós egységet tartalmaz, amelyek működése valószínűleg koordinált - a teljes tartomány aktív vagy inaktív állapotban van.
Domén szinten a kromatin szekvenciális pakolódása következtében a DNS lineáris mérete körülbelül 200-szorosára (700 nm) csökken.
- kromoszóma szint
Kromoszómális szinten a profázis kromoszóma kondenzációja a metafázis kromoszómába a hurokdomének tömörödésével megy végbe a nem hiszton fehérjék axiális kerete körül. Ezt a szuperspirálozást a sejtben lévő összes H1-molekula foszforilációja kíséri. Ennek eredményeként a metafázisú kromoszóma szorosan egymáshoz tömött szolenoid hurkokként ábrázolható, amelyek szoros spirálba tekerednek. Egy tipikus emberi kromoszóma akár 2600 hurkot is tartalmazhat. Egy ilyen szerkezet vastagsága eléri az 1400 nm-t (két kromatid), miközben a DNS-molekula 104-szeresére rövidül, azaz. 5 cm-es DNS-sel 5 μm-re feszített.
Kromoszóma funkciók
Az extrakromoszómális mechanizmusokkal kölcsönhatásban a kromoszómák biztosítják
- örökletes információk tárolása
- ezen információk felhasználásával a sejtes szervezet létrehozására és fenntartására
- az örökletes információk olvasásának szabályozása
- a genetikai anyag önmegkettőződése
- genetikai anyag átvitele az anyasejtből a lányba.
Bizonyíték van arra, hogy amikor a kromatin régió aktiválódik, pl. A transzkripció során először a H1 hiszton, majd a hisztonoktett távolodik el reverzibilisen. Ez a kromatin dekondenzációját okozza, egy 30 nanométeres kromatinszál szekvenciális átalakulását 10 nanométeres filamentummá, és további kibontakozását a szabad DNS régióiba, azaz. a nukleoszómális szerkezet elvesztése.
A megismerés ökológiája. Tudomány és felfedezés: Mi az élet? Ez a kérdés vált a genetika (a görög genetikos szóból – "születéssel, leszármazással kapcsolatos") – az élet keletkezésének tudománya – fejlődésének mozgatórugójává, amely már több mint 50 éve a DNS-molekulára fókuszál.
Mi az élet? Ez a kérdés vált a genetika (a görög genetikos szóból – "születéssel, leszármazással kapcsolatos") – az élet keletkezésének tudománya – fejlődésének mozgatórugójává, amely már több mint 50 éve a DNS-molekulára fókuszál.
A felfedezés, ami felforgatta a világot
– Most fedeztük fel az élet titkát! - így 1953. február 28-án Francis Crick és James Watson bejelentette, hogy felfedezték a DNS szerkezetét. Milyen újdonságot hozott az élettudományok számára? Korábban ismert volt, hogy a DNS egy nagy molekula, amelyben az élőlények szerkezetére és tulajdonságaira vonatkozó információkat a "négybetűs ábécé" segítségével rögzítik. De továbbra is tisztázatlan maradt, hogy ez az információ hogyan továbbadódik nemzedékről nemzedékre, és hogyan valósul meg ezekben a struktúrákban és tulajdonságokban, valamint hogy mi a DNS térbeli szerkezete.
A DNS szerkezetének megoldása segített a tudósoknak megérteni a DNS másolásának és materializálódásának mechanizmusait. A DNS két szálból áll, amelyek komplementerek (komplementerek) egymással. A DNS-másolás annak köszönhető, hogy minden kezdeti DNS-szálon egy másolatot adnak hozzá, mint a templáthoz. Tehát a DNS egy kettős hélixéből két teljesen azonos kettős hélix keletkezik, amelyek szükségesek a genetikai információ megőrzéséhez az anyasejt két leánysejtre való osztódása során. A mátrix elv a genetikai információ fokozatos materializálódásának is az alapja: az egyik DNS-szálon egy másik információs molekula - RNS - komplementer lánca képződik, amely viszont mátrixként szolgál a fehérjék szintéziséhez, mennyiségéhez és minőségéhez. amelyek közül az adott szervezet szerkezetétől és tulajdonságaitól függ.
Mennyire jelentős ez a felfedezés az élet titkának megértéséhez? Egyrészt a DNS szerkezetének ismerete nyilvánvalóan nem elegendő a „mi az élet?” kérdés megválaszolásához. De másrészt ez a felfedezés volt az, ami egy nagyon ősi és nagyon fontos kérdés a potenciál és a megnyilvánult kapcsolatáról - egy szervezet szerkezetére és tulajdonságaira vonatkozó információk magukkal a struktúrákkal és tulajdonságokkal való összekapcsolásának példáján. És nemcsak felvetette ezt a kérdést, hanem kulcsot is adott a válaszhoz. Ez a kulcs a mátrix elv, a komplementaritás elve.
Út a géntől a tulajdonságig
Mit jelent a tankönyv klasszikus mondata: "A DNS a genetikai információ hordozója"? Hogyan kapcsolódik a genetikai információ a DNS szerkezetéhez? Hogyan testesül meg az információ egy szervezet meghatározott tulajdonságaiban? Ha a genetikai információ kiindulási pontjának a DNS-szerkezetet vesszük, majd a szerkezeti modellt követjük, akkor a géntől a tulajdonságig vezető út így fog kinézni: a DNS-szekvenciában a szervezet minden tulajdonsága titkosítva van; egy adott gén lineáris szerkezete egyértelműen meghatározza a megfelelő fehérje lineáris szerkezetét, ami viszont egyértelműen meghatározza e fehérje szerepét egy adott tulajdonság kialakításában.
Más szavakkal: „A DNS RNS-t eredményez; Az RNS fehérjét szül, a fehérje pedig téged és engem” (F. Crick). Ha ez igaz, akkor egy adott tulajdonság megváltoztatásához (például egy genetikai gyökerű betegség gyógyításához) elegendő a DNS-szakasz megfelelő szekvenciáját létrehozni és kijavítani.
De ilyen egyszerű? Van-e elég tudás (bár vitathatatlanul helyes és szükséges!) a géntől a tulajdonságig vezető úton strukturális megfelelésekről ahhoz, hogy megértsük és reprodukáljuk ezt az utat?
A genetika legújabb eredményei azt mutatták, hogy ezek nem elegendőek. 2003-ban a Humán Genom Projekt keretében az emberi DNS (és sok más egyszerű és összetett organizmus) lineáris szerkezetét teljesen meghatározták. Ahogy az egyik tudós mondta: "az összes betűt, ami egy vastag könyvet alkot, elolvasták, most még meg lehetne érteni a szavakat és a jelentésüket."
Kiderült, hogy az emberben körülbelül 30 000 tulajdonképpeni gén (fehérjéket kódoló DNS-szakasz) található, és ez az összes DNS-nek csak 1-3%-a! Ugyanannyi gén az Arabidopsis taliana növényben és a gömbhalban. Ráadásul az emberi gének 99%-a egybeesik az egér génjeivel, vagyis az embernek mindössze 300 olyan génje van, amivel az egér nem rendelkezik. (Nehéz elképzelni, hogy nekünk és az egereknek a jelek 99%-a ugyanaz!)
Tovább tovább. Kiderült, hogy egy gén és egy fehérje között egyértelmű kapcsolat csak a baktériumokban létezik. Emberben pedig lehetőség van egy gén alapján sok fehérje kialakítására (az egy gén által kódolt különböző fehérjék maximális ismert száma ma 40 000!) És sok funkció megjelenése egy fehérjében. Kiderült, hogy az út a potenciáltól a megnyilvánultig, a genetikai információtól a tulajdonságig korántsem lineáris; hogy mindegyik tulajdonság számos gén és fehérjetermékeik közötti összetett kölcsönhatás eredménye; hogy maga a „gén” fogalma – kétértelműsége miatt – aligha szolgálhat ennek az útnak „kiindulópontjaként”.
A szerkezetek kölcsönhatása
Az emberi test 10 sejtből áll a 14. hatványig. Mindegyiknek pontosan ugyanaz a DNS-e, de jelentősen eltérnek egymástól alakjuk, méretük és feladataik tekintetében. Ennek a paradoxonnak a megoldása a genetikai információ szelektív leolvasásában rejlik. Minden sejtben csak azok a gének aktívak, amelyekre éppen szüksége van.
A szelektivitást speciális szabályozó gének biztosítják, amelyek lehetővé teszik vagy tiltják a DNS egy adott szakaszából származó információk kiolvasását. A génaktivitás a sejtmag terében lévő környezetétől is függ. Maga a gén vagy valamelyik szomszédjának mozgása által okozott környezeti változás megváltoztathatja annak aktivitását ("kikapcsolja" vagy "bekapcsolja" a gént).
Például az emberi genom sok potenciálisan veszélyes vírusgént és protoonkogént tartalmaz (amelyek egy sejt rákos degenerációját okozhatják). Hosszú ideig (és egész életükben) egészen békésen viselkedhetnek, sőt a sejt javára is dolgozhatnak, amíg saját maguk vagy a környezetből valaki mozgása agresszív potenciálokat nem tár fel ezekben a génekben. Szerencsére más mozgások is előfordulhatnak, amelyek megnyugtatják a "lázadót", vagy aktiválják a védekező mechanizmusokat.
Tehát a genetikai információ hordozója a gén szintjéről (a DNS egy meghatározott szakasza) egy epigenetikai (a latin "over", "over" szóból) a gének egymással és más struktúrákkal való interakciójának szintjére került. a sejtmag (a nem gén eredetű DNS és fehérjék 99%-a). Tegyük fel, hogy a tudomány megfejti ennek a kölcsönhatásnak a mechanizmusát. Ez az élet titkának felfedéséhez vezet? Az élet csak egy szerkezet? És ha nem, érdemes-e az élet misztériumának nyomát keresni a struktúrák kölcsönhatására korlátozódni?
Ki figyeli az őrt?
Hogyan lehet egyetlen sejtből 46 osztódás eredményeként nem 1014 sejtből álló alaktalan tömeget kapni, hanem mindannyiunkra nagyon jellemző testet? A sejtek egymás után megkétszereződve nemcsak önmagukban válnak különbözővé, hanem különböző testrészeket is alkotnak a megfelelő időés a megfelelő helyen. Mi szabályozza a sejtek időbeli és térbeli szerveződését?
Egy egész, amely minőségileg nagyobb, mint az alkotórészeinek-celláknak az egyszerű összege. És ez nem mond ellent annak, hogy a szervezet egy sejtből jön létre - a kérdés az, hogy ez a sejt miben testesíti meg az „egész akaratát”. Egy ilyen rendező tényező keresése a 20. század elején a morfogenetikus mező fogalmát eredményezte. Alapítója az orosz tudós, A.G. Gurvich. Amikor Gurvich a terepelmélettel foglalkozott, a DNS-molekulát a kromoszómák szerves részének tekintették, és nem tulajdonítottak neki nagy jelentőséget.
A tudós 1944-ben publikálta "A biológiai mező elmélete" című munkáját. Az idei év minden genetika számára döntő jelentőségűvé vált, és több évtizedre meghatározza fejlődésének útját. A tudósok fókuszában a DNS-molekula állt, mivel bebizonyosodott, hogy ő játszotta a vezető szerepet az örökletes információk továbbításában. 1953 már nem volt messze... Ennek eredményeként a tudomány minden figyelmét a DNS szerkezetére összpontosította, amelyet valójában elkezdtek azonosítani az általa hordozott információkkal, és a biológiai mező elméletét nem tartották nagy becsben. . De a kutatás ezen a területen folytatódott, és az évek során az élet titkainak megismerésének két útja párhuzamosan haladt ...
Az új évezredre való átmenet megváltoztatta az élettudományok erőviszonyait. Egyre több tudós jut arra a következtetésre, hogy az élők tudásának szerkezeti kulcsa szükséges, de nem elégséges; hogy a különböző megközelítések nem zárják ki egymást, hanem a tudományos kutatás egységes útját képezik; hogy lényegében a strukturális megközelítés és a térelmélet kiegészítik egymást.
Emlékezzünk: a DNS-láncok komplementaritásának feltételezése volt a kulcsa annak szerkezetének megfejtéséhez, és maga az 1953-as felfedezés is a különböző tudományterületek képviselői – fizikusok, kémikusok, biológusok – erőfeszítéseinek komplementaritása révén vált lehetővé. Talán az új évezred egységes tudománya nemcsak a különböző tudományos (például szerkezeti és területi) megközelítéseket fogja végre összhangba hozni, hanem az élet titkainak „tudománytalan” megismerésének gyümölcsei felé is fordul – az ezeréves bölcsesség felé. az emberiség - "tudománytalan", mivel ez az út azokban az időkben gyökerezik, amikor egyáltalán nem volt tudomány.
Az ősi bölcsesség forrásaihoz fordulva megadhatja a tudománynak a kulcsokat az ajtóhoz, amely mögött a titok rejtőzik. De hogy ez megtörténjen, szóval különböző utak a tudásnak valahol "metszenie kell". Az egyik ilyen „kereszteződés” lehet a formaképző mező (biológiai, morfogenetikai, információs) fogalma, amely a modern tudomány alapján fejlődött ki, és az ókori forrásokban tárgyalt jelenségekre nyúlik vissza.
Utóbbiak azt mondják, hogy egy személy több testből vagy princípiumból áll, amelyek nem különálló, független részek, hanem áthatolnak és kölcsönösen formálják egymást; hogy a látható, sűrű, fizikai test karmester, finomabb testek hordozója, amelyek segítségével megnyilvánulnak a fizikai világban és kölcsönhatásba lépnek vele; hogy az "összeállítást" - a fizikai test kialakítását a fizikai anyag elemeiből - ezek közül a testek "legsűrűbbje", az asztráltest prototípusa (terv, mátrix) irányítja.
A modern tudomány tudja, hogy minden újonnan képződött sejt egyéni „utasítás” (aktív, vagy bekapcsolt gének) szerint vesz részt egy szervezet kialakításában, és ennek a sejtnek a szülője és leszármazottja teljesen eltérő „utasításokkal” rendelkezhet. De hogy mi és hogyan koordinálja majd az egyes "programok" váltását több milliárd sejt fejlesztésére, az még nem világos.
A biológiai térelmélet feltételezi, hogy az egyetértés az egész függvénye, amely maga a mező, mátrix vagy modell; hogy minden újonnan képződött sejt a saját genetikai apparátusa segítségével a szervezet fejlődésének egyetlen "tervéhez" kapcsolódik, onnan kapja az egyedi "utasításokat" és azokat saját viselkedési programja keretein belül valósítja meg.
Kiderült, hogy a sejt genetikai berendezése legalább három funkcionális blokkból áll: egy vevő „antennából”, egy „vezérlőpultból” a génaktivitáshoz és egy „végrehajtó” részből - specifikus fehérjék kialakulásáért felelős génekből. Emlékezzünk vissza, hogy a gének az összes sejt DNS-ének csak 1-2%-át teszik ki. A DNS fennmaradó 98-99%-ában már találtak a második „kontrollblokkhoz” kapcsolódó struktúrákat. És mi játssza az "antenna" szerepét? Hol történik a "két világ találkozása" - az információs mező és a genetikai struktúrák, amelyek ezt az információt megtestesítik egy adott fizikai testben?
Miért nem feltételezzük, hogy a DNS egy antenna szerepét is betölti, amely képes felvenni, átalakítani és továbbítani a hullámjellegű jeleket? Az "életmolekula" spirális szerkezete (sok műszaki antenna spirál alakú) és tulajdonságai, mint az elektromos áram vezethetősége, a hosszirányú rezgések rezonáns gerjesztésének lehetősége rádióhullámok hatására, valamint mint a lézerfény előállításának képessége az előzetes "szivattyúzás" után.
Ha a DNS képes arra, hogy olyan információt fogadjon, amely biztosítja a sejtek létfontosságú tevékenységét, akkor nem kell folyamatosan tárolnia ezt az információt a szerkezetében. Ahogy például az emberi agynak ahhoz, hogy sikeresen irányítsa a test életfenntartó rendszereit, nem kell az elme "fogadójának" lennie, hanem elegendő közvetítő szerepet játszani a tudat és a test: a tudat síkjáról érzékeli az információt és "lefordítja" a testkontroll nyelvére.
És világos, hogy a fizikai test miért szenved a DNS-szerkezet (vagy az agyi struktúrák) károsodása esetén. Hiszen mindenki tudja, hogy a TV legalább egy részének meghibásodása esetén a képernyőn látható kép erősen torzul, és ha a TV-t megfosztják az antennától vagy kikapcsolják a hálózatról, akkor semmi sem jelenik meg a képernyőn. képernyő egyáltalán.
A DNS a kapcsolat a fizikai test "modellje" és konkrét megtestesülése között. Az agy közvetítő az elme és a test között. Az értelem összekapcsolja az életet és megnyilvánulási formáját, és lehetővé teszi a formába zárt élet számára, hogy megismerje önmagát. Ennek a csodálatos eszköznek a segítségével az embernek lehetősége nyílik arra, hogy tanulmányozza az őt körülvevő világot, és megtalálja benne a kulcsokat belső világa megismeréséhez. Így születik egy egységes út, amely az élet titkának megismeréséhez vezet. Mert az ember a legnagyobb rejtély – a föld és az ég kapcsolatának titka. közzétett
P.S. És ne feledje, pusztán a fogyasztás megváltoztatásával - együtt megváltoztatjuk a világot! © econet
Csatlakozzon hozzánk a
Miután 1953-ban felfedezték egy ilyen anyag, például a DNS molekuláris szerveződésének elvét, a molekuláris biológia fejlődésnek indult. Továbbá a kutatás során a tudósok rájöttek a DNS rekombinációjára, összetételére és emberi genomunk működésére.
A legösszetettebb folyamatok molekuláris szinten zajlanak nap mint nap. Hogyan épül fel egy DNS-molekula, miből áll? És milyen szerepet játszanak a DNS-molekulák a sejtben? Beszéljünk részletesen a kettős láncon belül zajló folyamatokról.
Mi az örökletes információ?
Szóval hol kezdődött az egész? Még 1868-ban baktériumok magjában találták meg őket. 1928-ban pedig N. Kolcov azt az elméletet terjesztette elő, hogy a DNS-ben van az, hogy az élő szervezetről szóló összes genetikai információ titkosítva van. Aztán J. Watson és F. Crick 1953-ban megtalálta a ma ismert DNS-spirál modelljét, amiért méltán kaptak elismerést és díjat - Nóbel díj.
Egyébként mi az a DNS? Ez az anyag 2 egyesített szálból, pontosabban spirálokból áll. Egy ilyen lánc bizonyos információkat tartalmazó szakaszát génnek nevezzük.

A DNS tárol minden információt arról, hogy milyen fehérjék képződnek és milyen sorrendben. A DNS-makromolekula hihetetlenül terjedelmes információ anyagi hordozója, amelyet az egyes építőelemek - nukleotidok - szigorú sorrendje rögzít. Összesen 4 nukleotid van, ezek kémiailag és geometriailag kiegészítik egymást. A tudományban a komplementaritásnak vagy komplementaritásnak ezt az elvét később ismertetjük. Ez a szabály kulcsszerepet játszik a genetikai információ kódolásában és dekódolásában.
Mivel a DNS-szál hihetetlenül hosszú, ebben a szekvenciában nincsenek ismétlések. Minden élőlénynek megvan a maga egyedi DNS-szála.
DNS funkciók
A funkciók közé tartozik az örökletes információk tárolása és utódoknak való továbbítása. E funkció nélkül a faj genomja évezredekig nem maradhatott meg és nem fejleszthető. Azok a szervezetek, amelyek jelentős génmutáción estek át, nagyobb valószínűséggel maradnak életben, vagy elveszítik az utódnemzés képességét. Így történik a természetes védekezés a fajok elfajulása ellen.

Egy másik lényeges funkció a tárolt információk megvalósítása. Egy sejt nem tud egyetlen létfontosságú fehérjét előállítani a kettős láncban tárolt utasítások nélkül.
Nukleinsav összetétel
Ma már megbízhatóan ismert, hogy maguk a nukleotidok - a DNS építőkövei - miből állnak. 3 anyagból állnak:
- Ortofoszforsav.
- Nitrózus alap. Pirimidin bázisok - amelyeknek csak egy gyűrűjük van. Ezek közé tartozik a timin és a citozin. Purin bázisok, amelyek 2 gyűrűt tartalmaznak. Ezek a guanin és az adenin.
- Szacharóz. A DNS részeként - dezoxiribóz, RNS-ben - ribóz.
A nukleotidok száma mindig megegyezik a nitrogéntartalmú bázisok számával. Speciális laboratóriumokban egy nukleotidot hasítanak, és nitrogénbázist izolálnak belőle. Így tanulmányozzák ezeknek a nukleotidoknak az egyedi tulajdonságait és a bennük lévő lehetséges mutációkat.
Az örökletes információk szerveződési szintjei
A szerveződésnek 3 szintje van: gén, kromoszómális és genomiális. Az új fehérje szintéziséhez szükséges összes információ a lánc egy kis szakaszában - a génben - található. Vagyis a gént az információkódolás legalacsonyabb és legegyszerűbb szintjének tekintik.

A gének pedig kromoszómákká állnak össze. Az örökletes anyag hordozójának ilyen szervezetének köszönhetően a karaktercsoportok bizonyos törvények szerint váltakoznak, és egyik generációról a másikra továbbadódnak. Meg kell jegyezni, hogy hihetetlenül sok gén található a szervezetben, de az információ nem vész el még akkor sem, ha többszörös rekombinációra kerül sor.
Többféle gén létezik:
- funkcionális rendeltetésük szerint 2 típusa van: szerkezeti és szabályozó szekvenciák;
- a sejtben lezajló folyamatokra gyakorolt hatás szerint megkülönböztetik: szupervitalis, letális, feltételesen letális gének, valamint mutátor és antimutátor gének.
A gének a kromoszóma mentén lineáris sorrendben helyezkednek el. A kromoszómákban az információ nem véletlenszerűen fókuszál, van egy bizonyos sorrend. Van még egy térkép is, amely a gének helyzetét vagy lokuszát mutatja. Ismeretes például, hogy a 18-as kromoszóma a gyermek szemének színére vonatkozó adatokat kódolja.
Mi az a genom? Ez a neve a test sejtjében található nukleotidszekvenciák teljes készletének. A genom az egész fajt jellemzi, nem egy egyedet.
Mi az emberi genetikai kód?
A tény az, hogy az összes hatalmas potenciál az emberi fejlődés már a fogantatás időszakában lefektették. Minden örökletes információ, amely a zigóta fejlődéséhez és a gyermek születés utáni növekedéséhez szükséges, a génekben kódolva van. A DNS részei az örökletes információ legalapvetőbb hordozói.
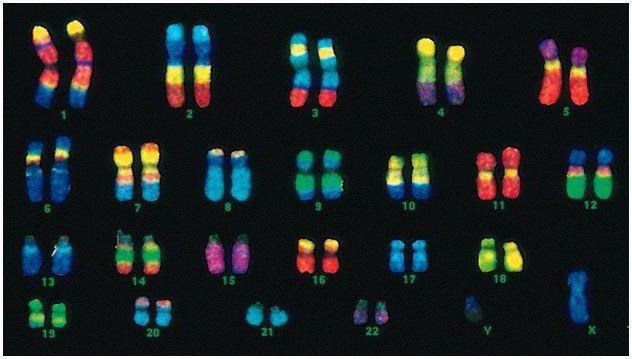
Egy személynek 46 kromoszómája vagy 22 szomatikus párja van, plusz egy nemet meghatározó kromoszóma mindegyik szülőtől. Ez a diploid kromoszómakészlet az ember teljes fizikai megjelenését, szellemi és fizikai képességeit, valamint betegségekre való hajlamát kódolja. Szomatikus kromoszómák külsőleg megkülönböztethetetlenek, de más-más információt hordoznak, hiszen az egyik az apától, a másik az anyától származik.
A férfi kód eltér a női kódtól az utolsó kromoszómapárban - XY. A női diploid halmaz az utolsó pár, a XX. A férfiak egy X-kromoszómát kapnak biológiai anyjuktól, majd azt továbbadják lányaiknak. A nemi Y kromoszóma a fiakra kerül.
Az emberi kromoszómák mérete nagyon eltérő. Például a legkisebb kromoszómapár a # 17. És a legnagyobb pár az 1 és a 3.
A kettős hélix átmérője emberben mindössze 2 nm. A DNS olyan szorosan csavarodott, hogy belefér a sejt kis magjába, bár felcsavarva akár 2 méter hosszú is lehet. A hélix hossza több száz millió nukleotid.
Hogyan továbbítható a genetikai kód?
Tehát milyen szerepet játszanak a DNS-molekulák a sejtben az osztódás során? A gének - az örökletes információ hordozói - a test minden sejtjében megtalálhatók. Sok lény, hogy átadja kódját egy leányszervezetnek, két azonos spirálra osztja DNS-ét. Ezt replikációnak nevezik. A replikáció során a DNS feltekercselődik, és speciális "gépek" egészítik ki az egyes szálakat. A genetikai spirál kettéágazása után a sejtmag és az összes organellum osztódni kezd, majd az egész sejt.
De egy személynek más a génátviteli folyamata - szexuális. Az apa és az anya vonásai keverednek, az új genetikai kód mindkét szülőtől származó információkat tartalmaz.
Az örökletes információk tárolása és továbbítása a DNS-hélix összetett szerveződésének köszönhetően lehetséges. Végül is, mint mondtuk, a fehérjék szerkezetét gének kódolják. A fogantatáskor létrejött kód egész élete során önmagát másolja. A kariotípus (személyes kromoszómakészlet) nem változik a szervsejtek megújulása során. Az információátadás szexuális ivarsejtek - férfi és női - segítségével történik.
Csak az egy RNS-szálat tartalmazó vírusok nem képesek információikat továbbítani utódaiknak. Ezért szaporodásukhoz emberi vagy állati sejtekre van szükségük.
Örökletes információk realizálása
A sejtmagban, fontos folyamatokat... A kromoszómákban rögzített összes információt felhasználják aminosavakból fehérjék felépítésére. De a DNS-szál soha nem hagyja el a sejtmagot, ezért itt egy másik fontos vegyület = RNS segítségére van szükség. Ez az RNS, amely képes áthatolni a nukleáris membránon és kölcsönhatásba lépni a DNS-száljal.
A DNS és 3 típusú RNS kölcsönhatása révén minden kódolt információ megvalósul. Milyen szinten áll az örökletes információ megvalósítása? Minden kölcsönhatás nukleotid szinten megy végbe. A hírvivő RNS lemásolja a DNS-lánc egy részét, és ezt a másolatot a riboszómába viszi. Itt kezdődik egy új molekula szintézise nukleotidokból.
Annak érdekében, hogy az mRNS lemásolja a szál kívánt részét, a hélixet kibontják, majd az újrakódolási folyamat befejezése után újra helyreállítják. Ráadásul ez a folyamat egy kromoszóma két oldalán egyidejűleg is végbemehet.
A komplementaritás elve
4 nukleotidból állnak - adenin (A), guanin (G), citozin (C), timin (T). A komplementaritási szabály szerint hidrogénkötésekkel kapcsolódnak össze. E. Chargaff munkái segítettek ennek a szabálynak a megállapításában, mivel a tudós bizonyos mintákat észlelt ezen anyagok viselkedésében. E. Chargaff felfedezte, hogy az adenin és a timin mólaránya egy. És ugyanígy a guanin és a citozin aránya mindig eggyel egyenlő.
Munkássága alapján a genetika szabályt alkotott a nukleotidok kölcsönhatására. A komplementaritás szabálya szerint az adenin csak a timinnel, a guanin pedig a citozinnal kombinálódik. A hélix dekódolása és egy új fehérje szintézise során a riboszómában egy ilyen váltakozó szabály segít gyorsan megtalálni esszenciális aminosav amely a transzport RNS-hez kapcsolódik.
Az RNS és típusai
Mi az örökletes információ? nukleotidok a DNS kettős szálában. Mi az az RNS? Mi a munkája? Az RNS, vagyis a ribonukleinsav segít a DNS-ből információk kinyerésében, dekódolásában és a komplementaritás elve alapján a sejtekhez szükséges fehérjék létrehozásában.
Összesen 3 típusú RNS-t izolálnak. Mindegyik szigorúan betölti funkcióját.
- Információs (mRNS), vagy mátrixnak hívják. Egyenesen a sejt közepébe, a sejtmagba kerül. Az egyik kromoszómában megtalálja a fehérje felépítéséhez szükséges genetikai anyagot, és lemásolja a kettős lánc egyik oldalát. A másolás ismét a komplementaritás elve szerint történik.
- Szállítás Egy kis molekula, amelynek egyik oldalán nukleotid dekóderek, másik oldalán pedig az alapkódnak megfelelő aminosavak találhatók. A tRNS feladata, hogy eljuttassa a „műhelybe”, vagyis a riboszómába, ahol szintetizálja a szükséges aminosavat.
- rRNS - riboszómális. Szabályozza a termelődő fehérje mennyiségét. 2 részből áll - aminosav és peptid hely.
Az egyetlen különbség a dekódolásban az, hogy az RNS nem tartalmaz timint. A timin helyett uracil van. De aztán a fehérjeszintézis folyamatában a tRNS-sel még mindig helyesen állítja be az összes aminosavat. Ha az információ dekódolása során hiba lép fel, akkor mutáció lép fel.
A sérült DNS-molekula javítása
A sérült kettős szál javításának folyamatát javításnak nevezzük. A javítási folyamat során a sérült géneket eltávolítják.

Ezután a kívánt elemsort pontosan reprodukálják, és visszavágják a lánc ugyanarra a helyére, ahonnan kivonták. Mindez a speciálisnak köszönhetően történik vegyszerek- enzimek.
Miért fordulnak elő mutációk?
Miért kezdenek el egyes gének mutálódni, és nem teljesítik funkciójukat – létfontosságú örökletes információkat tárolnak? Ennek oka egy dekódolási hiba. Például, ha az adenint véletlenül timinre cserélik.
Vannak kromoszómális és genomi mutációk is. A kromoszómális mutációk akkor fordulnak elő, amikor az örökletes információ egy része kiesik, megkettőződik, vagy akár átkerül és beépül egy másik kromoszómába.

A genomi mutációk a legsúlyosabbak. Oka a kromoszómák számának megváltozása. Vagyis amikor egy pár - diploid halmaz helyett egy triploid halmaz van jelen a kariotípusban.
A triploid mutáció leghíresebb példája a Down-szindróma, amelyben a személyes kromoszómakészlet 47. Az ilyen gyerekeknél a 21. pár helyett 3 kromoszóma képződik.
Egy mutáció, például a poliploidia is ismert. A poliploidia azonban csak növényekben fordul elő.
A DNS-molekula két szálból áll, amelyek kettős hélixet alkotnak. Felépítését először Francis Crick és James Watson fejtette meg 1953-ban.
Eleinte egy DNS-molekula, amely egy pár nukleotidláncból áll, amelyek egymás köré csavarodtak, kérdéseket vetett fel, hogy miért pont ilyen alakú. A tudósok ezt a jelenséget komplementaritásnak nevezték, ami azt jelenti, hogy csak bizonyos nukleotidok helyezkedhetnek el egymással szemben lévő szálaiban. Például az adenin mindig a timinnel, a guanin pedig a citozinnal szemben áll. A DNS-molekula ezen nukleotidjait komplementernek nevezzük.
Ezt sematikusan a következőképpen ábrázoljuk:
T-A
C-G
Ezek a párok kémiai nukleotidkötést alkotnak, amely meghatározza az aminosavak elrendezésének sorrendjét. Az első esetben valamivel gyengébb. C és G között erősebb a kapcsolat. A nem komplementer nukleotidok nem párosulnak egymással.
A szerkezetről
Tehát a DNS-molekula szerkezete különleges. Okkal van ilyen alakja: tény, hogy a nukleotidok száma nagyon nagy, és sok hely kell a hosszú láncok befogadásához. Ez az oka annak, hogy a spirálcsavarás a láncok velejárója. Ezt a jelenséget spiralizációnak nevezik, ez lehetővé teszi a filamentumok körülbelül öt-hatszoros rövidülését.
A szervezet egyes ilyen molekulákat nagyon aktívan, másokat ritkán használ. Ez utóbbiak a spiralizálás mellett olyan "kompakt csomagoláson" is átesnek, mint a szupertekercselés. És akkor a DNS-molekula hossza 25-30-szor csökken.

Mi a molekula "csomagolása"?
A szuperspirálozás folyamatában hisztonfehérjék vesznek részt. Olyan szerkezetűek és megjelenésűek, mint egy menetes orsó vagy rúd. Spiralizált szálak vannak rájuk tekercselve, amelyek azonnal "kompaktra tömörülnek", és kevés helyet foglalnak el. Amikor szükségessé válik az egyik vagy másik szál használata, azt például egy hisztonfehérje tekercséről letekerik, és a spirál két párhuzamos láncra tekerődik. Amikor a DNS-molekula ilyen állapotban van, a szükséges genetikai adatok kiolvashatók belőle. Van azonban egy feltétel. Információszerzés csak akkor lehetséges, ha a DNS-molekula szerkezete nem csavarodott. Az olvasásra rendelkezésre álló kromoszómákat euchromatinoknak nevezzük, és ha szuperszipiralizáltak, akkor ezek már heterokromatinok.
Nukleinsavak
A nukleinsavak a fehérjékhez hasonlóan biopolimerek. A fő funkció az örökletes (genetikai) információk tárolása, megvalósítása és továbbítása. Két típusuk van: DNS és RNS (dezoxiribonukleinsav és ribonukleinsav). A bennük lévő monomerek nukleotidok, amelyek mindegyike tartalmaz egy foszforsav-maradékot, egy öt szénatomos cukrot (dezoxiribóz / ribóz) és egy nitrogénbázist. A DNS-kód 4 típusú nukleotidot tartalmaz - adenin (A) / guanin (G) / citozin (C) / timin (T). A bennük lévő nitrogénbázisban különböznek.
Egy DNS-molekulában a nukleotidok száma óriási lehet – több ezertől tíz- és százmillióig. Az ilyen óriásmolekulákat elektronmikroszkóppal lehet megnézni. Ebben az esetben látható lesz a polinukleotid szálak kettős szála, amelyeket a nukleotidok nitrogéntartalmú bázisainak hidrogénkötései kapcsolnak össze.

Kutatás
A kutatás során a tudósok azt találták, hogy a DNS-molekulák típusai a különböző élő szervezetekben eltérőek. Azt is megállapították, hogy az egyláncú guanin csak a citozinhoz, a timin pedig az adeninhez kötődhet. Az egyik szál nukleotidjainak elrendezése szigorúan megfelel a párhuzamosnak. A polinukleotidok ezen komplementaritása miatt a DNS-molekula képes megkettőzésre és önreprodukcióra. De először a komplementer láncok szétválnak speciális enzimek hatására, amelyek elpusztítják a páros nukleotidokat, majd mindegyikben megkezdődik a hiányzó lánc szintézise. Ez annak köszönhető, hogy a egy nagy szám szabad nukleotidok minden sejtben. Ennek eredményeként a "szülőmolekula" helyett két "leány" molekula jön létre, amelyek összetételében és szerkezetében azonosak, és a DNS-kód lesz az eredeti. Ez a folyamat a sejtosztódás előfutára. Biztosítja az összes örökletes adat átvitelét az anyasejtekből a leánysejtekbe, valamint az összes következő generációba.
Hogyan olvasható a génkód?
Ma már nemcsak a DNS-molekula tömegét számolják ki, hanem olyan összetettebb adatokat is meg lehet találni, amelyek korábban nem álltak a tudósok rendelkezésére. Például információkat olvashat arról, hogy a test hogyan használja a saját sejtjét. Természetesen eleinte ezek az információk kódolva vannak, és valamilyen mátrix formájúak, ezért egy speciális hordozóra, az RNS-re kell szállítani. A ribonukleinsav a sejtmag membránján keresztül képes behatolni a sejtbe, és be tudja olvasni a benne található kódolt információkat. Az RNS tehát rejtett adatok hordozója a sejtmagból a sejtbe, és abban különbözik a DNS-től, hogy dezoxiribóz helyett ribózt, timint helyett uracilt tartalmaz. Ezenkívül az RNS egyszálú.

RNS szintézis
A DNS mélyelemzése kimutatta, hogy miután az RNS elhagyja a sejtmagot, bejut a citoplazmába, ahol templátként beépülhet a riboszómákba (speciális enzimrendszerekbe). A kapott információk alapján szintetizálni tudják a fehérje aminosavak megfelelő szekvenciáját. A riboszóma a triplet kódból tanulja meg, hogy milyen szerves vegyületnek kell kapcsolódnia a kialakuló fehérjelánchoz. Minden aminosavnak megvan a saját specifikus triplettje, amely kódolja azt.
A lánc kialakulása után sajátos térbeli formát nyer, és olyan fehérjévé alakul, amely képes ellátni hormonális, építő, enzimatikus és egyéb funkcióit. Bármely szervezet számára ez egy géntermék. Ez határozza meg a gének mindenféle tulajdonságát, tulajdonságait és megnyilvánulásait.

Gének
Mindenekelőtt szekvenálási eljárásokat dolgoztak ki annak érdekében, hogy információt szerezzenek arról, hogy egy DNS-molekula szerkezete hány génből áll. És bár a kutatás lehetővé tette a tudósok számára, hogy nagy előrelépést érjenek el ebben a kérdésben, még nem lehet tudni pontos számukat.
Néhány évvel ezelőtt azt feltételezték, hogy a DNS-molekulák körülbelül 100 ezer gént tartalmaznak. Kicsit később ez a szám 80 ezerre csökkent, és 1998-ban a genetikusok bejelentették, hogy egyetlen DNS-ben mindössze 50 ezer gén található, ami a DNS teljes hosszának mindössze 3%-a. A genetikusok legújabb következtetései azonban megdöbbentek. Most azt állítják, hogy a genom 25-40 ezer ilyen egységet tartalmaz. Kiderült, hogy a kromoszómális DNS mindössze 1,5%-a felelős a fehérjék kódolásáért.
A kutatás ezzel nem állt meg. Egy párhuzamos géntechnológiai szakemberekből álló csapat megállapította, hogy egy molekulában pontosan 32 ezer a gének száma. Amint látja, még mindig lehetetlen végleges választ kapni. Túl sok az ellentmondás. Minden kutató csak a saját eredményeire támaszkodik.
Történt evolúció?
Annak ellenére, hogy nincs bizonyíték a molekula evolúciójára (mivel a DNS-molekula szerkezete törékeny és kis méretű), a tudósok egy javaslatot tettek. Laboratóriumi adatok alapján a következő tartalom egy változatát hangoztatták: megjelenésének kezdeti szakaszában a molekula egy egyszerű önreplikáló peptidnek tűnt, amely akár 32, az ősi óceánokban található aminosavat tartalmazott.
Az önreplikáció után a természetes szelekció erőinek köszönhetően a molekulák képesek voltak megvédeni magukat a külső elemek befolyásától. Kezdtek tovább élni és nagy számban szaporodtak. A lipidhólyagban talált molekulák minden esélyt megkaptak az önszaporodásra. Az egymást követő ciklusok eredményeként a lipidbuborékok sejtmembránok formáját öltötték, és csak ezután - minden ismert részecskék... Meg kell jegyezni, hogy ma a DNS-molekula bármely része összetett és egyértelműen működő szerkezet, amelynek minden jellemzőjét a tudósok még nem tanulmányozták teljesen.

Modern világ
A közelmúltban izraeli tudósok olyan számítógépet fejlesztettek ki, amely másodpercenként billió műveletet képes végrehajtani. Ma ez a leggyorsabb autó a Földön. Az egész titok az, hogy az innovatív eszközt a DNS hajtja. A professzorok szerint rövid távon az ilyen számítógépek akár áramot is termelhetnek majd.
Egy évvel ezelőtt a rehovoti (Izrael) Weizmann Intézet szakemberei egy molekulákból és enzimekből álló, programozható molekuláris számítógép létrehozását jelentették be. Szilícium mikrochipeket cseréltek rájuk. A csapat mostanra még halad előre. Most már csak egyetlen DNS-molekula képes ellátni a számítógépet a szükséges adatokkal és a szükséges üzemanyaggal.
A biokémiai „nanoszámítógépek” nem fikció, a természetben már léteznek, és minden élőlényben megnyilvánulnak. De gyakran nem az emberek irányítják őket. Egy személy még nem tudja megoperálni egyetlen növény genomját sem azért, hogy kiszámolja mondjuk a "pi" számot.
Az a gondolat, hogy a DNS-t adatok tárolására/feldolgozására használják, először 1994-ben merült fel a tudósok fényes elméjében. Ekkor használtak egy molekulát egy egyszerű matematikai feladat megoldására. Azóta számos kutatócsoport javasolta a DNS-számítógépekkel kapcsolatos különféle projekteket. De itt minden próbálkozás csak az energiamolekulán alapult. Egy ilyen számítógépet szabad szemmel nem látni, úgy néz ki, mint egy átlátszó vízoldat egy kémcsőben. Nincsenek benne mechanikus alkatrészek, csak több billió biomolekuláris eszköz – és ez csak egy csepp folyadékban van!
Emberi DNS
Hogy milyen emberi DNS-ről van szó, 1953-ban ismerték meg az emberek, amikor a tudósok először tudták bemutatni a világnak a DNS kétszálú modelljét. Ezért Kirk és Watson Nobel-díjat kapott, mivel ez a felfedezés a 20. században alapvetővé vált.
Idővel persze bebizonyították, hogy egy strukturált emberi molekula nem csak úgy nézhet ki, mint a javasolt változatban. A DNS részletesebb elemzése után felfedezték a Z- A-, B- és balkezes formáját. Az A- forma gyakran kivétel, mivel csak nedvességhiány esetén képződik. De ez csak laboratóriumi vizsgálatokban lehetséges, a természetes környezet számára ez abnormális, élő sejtben ilyen folyamat nem fordulhat elő.
A B-forma klasszikus, dupla jobbos láncként ismert, de a Z-forma nem csak az ellenkező irányba, balra csavarodott, hanem cikkcakkosabb megjelenésű is. A tudósok azonosították a G-quadruplex formát is. A szerkezetében nem 2, hanem 4 szál van. A genetikusok szerint ez a forma azokon a területeken fordul elő, ahol túl sok guanin van.

Mesterséges DNS
Már ma is létezik mesterséges DNS, amely a valódi másolata; tökéletesen megismétli a természetes kettős hélix szerkezetét. De a primordiális polinukleotidtól eltérően a mesterségesben csak két további nukleotid található.
Mivel a szinkronizálás a valódi DNS különféle vizsgálatai során szerzett információk alapján jött létre, másolható, önreplikálható és fejleszthető is. A szakértők körülbelül 20 éve dolgoznak egy ilyen mesterséges molekula létrehozásán. Az eredmény egy csodálatos találmány, amely a genetikai kódot ugyanúgy tudja használni, mint a természetes DNS-t.
A négy elérhető nitrogénbázishoz a genetika további kettőt adott, amelyeket a természetes bázisok kémiai módosításával hoztak létre. A természetes DNS-sel ellentétben a mesterséges DNS meglehetősen rövid. Csak 81 bázispárt tartalmaz. Ugyanakkor szaporodik és fejlődik is.
A kapott molekula replikációja mesterségesen, a polimeráz láncreakciónak köszönhetően megy végbe, de ez egyelőre nem önállóan, hanem tudósok közreműködésével történik. Önállóan hozzáadják a szükséges enzimeket az említett DNS-hez, speciálisan elkészített folyékony közegbe helyezve.
Végeredmény
A DNS-fejlődés folyamatát és végeredményét különféle tényezők, például mutációk befolyásolhatják. Ez szükségessé teszi az anyagminták tanulmányozását, hogy az elemzések eredménye megbízható és megbízható legyen. Ilyen például az apasági teszt. De jó hír, hogy ritkák az olyan események, mint a mutáció. Ennek ellenére az anyagmintákat mindig újraellenőrzik annak érdekében, hogy az elemzés alapján pontosabb információkat kapjanak.

Növényi DNS
A magas szekvenálási technológiáknak (HTS) köszönhetően forradalom történt a genomika területén - a növényekből DNS kinyerése is lehetséges. Természetesen a növényi anyagokból jó minőségű DNS molekulatömeg megszerzése nehézségeket okoz a nagyszámú DNS mitokondriális és kloroplasztisz kópiák miatt, valamint magas szint poliszacharidok és fenolos vegyületek. A vizsgált szerkezet elkülönítésére ebben az esetben többféle módszert alkalmazunk.
Hidrogénkötés a DNS-ben
A DNS-molekulában lévő hidrogénkötés felelős a pozitív töltésű hidrogénatom között létrejövő elektromágneses vonzásért, amely egy elektronegatív atomhoz kapcsolódik. Ez a dipólus kölcsönhatás nem felel meg a kritériumnak kémiai kötés... De megvalósítható intermolekulárisan vagy a molekula különböző részein, azaz intramolekulárisan.

A hidrogénatom az elektronegatív atomhoz kapcsolódik, amely ennek a kötésnek a donorja. Az elektronegatív atom lehet nitrogén, fluor, oxigén. Ez - decentralizáció révén - elektronfelhőt vonz magához a hidrogénmagból és a hidrogénatomot (részben) pozitív töltésűvé teszi. Mivel a H mérete kicsi más molekulákhoz és atomokhoz képest, a töltés is kicsi.
DNS dekódolás
A DNS-molekula dekódolása előtt a tudósok először rengeteg sejtet vesznek fel. A legpontosabb és legsikeresebb munkához körülbelül egy millióra van szükségük. A vizsgálat során kapott eredményeket folyamatosan összehasonlítjuk és rögzítjük. A genomdekódolás ma már nem ritkaság, hanem megfizethető eljárás.

Természetesen egy sejt genomjának megfejtése nem célszerű feladat. Az ilyen vizsgálatok során nyert adatok nem érdeklik a tudósokat. De fontos megérteni, hogy az összes jelenleg létező dekódolási módszer összetettsége ellenére nem elég hatékony. Csak a DNS 40-70%-át teszik lehetővé.
A harvardi professzorok azonban nemrégiben bejelentettek egy módot a genom 90%-ának megfejtésére. A technika azon alapul, hogy az izolált sejtekhez primer molekulákat adnak, amelyek segítségével megindul a DNS replikációja. De még ez a módszer sem tekinthető sikeresnek, még véglegesíteni kell, mielőtt a tudományban nyíltan alkalmazzák.
DNS (dezoxiribonukleinsav) utal (az RNS-sel együtt), amelyek polimerek, vagy inkább polinukleotidok (monomer - nukleotid).
A DNS felelős a genetikai kód tárolásáért és továbbításáért a sejtosztódás során. A DNS-molekulákon keresztül valósul meg az öröklődés és a változékonyság. Minden típusú RNS szintetizálódik a DNS-en. Ezenkívül a különböző típusú RNS-ek együttesen biztosítják a sejtfehérjék szintézisét, azaz genetikai információkat valósítanak meg.
Az eukarióta sejtekben a DNS túlnyomó része a sejtmagban található, ahol speciális fehérjékkel komplexeket képeznek, ami kromoszómák képződését eredményezi. A prokarióta sejtekben egy nagy cirkuláris (vagy lineáris) DNS-molekula található (szintén fehérjékkel komplexben). Ezenkívül az eukarióta sejteknek saját DNS-ük van a mitokondriumokban és a kloroplasztiszokban.
A DNS esetében minden nukleotid 1) nitrogénbázisból áll, amely lehet adenin, guanin, citozin vagy timin, 2) dezoxiribóz, 3) foszforsav.
A DNS-szál nukleotidsorrendje határozza meg a molekula elsődleges szerkezetét. A DNS-t a molekula másodlagos szerkezete jellemzi, kettős hélix formájában (leggyakrabban jobbkezes). Ebben az esetben két DNS-szálat a komplementer nitrogénbázisok között létrejövő hidrogénkötések kötnek össze.
Az adenin a timin, a guanin pedig a citozin komplementere. Két hidrogénkötés jön létre az adenin és a timin között, három pedig a guanin és a citozin között. Így a guanin és a citozin egy kicsit erősebben kapcsolódik egymáshoz (bár a hidrogénkötések elvileg gyengék). A kötések számát a molekulák szerkezeti sajátosságai határozzák meg.
Az adenin és a guanin purinok, és két gyűrűből állnak. A timin és a citozin pirimidinbázisokra utalnak, amelyek egy gyűrűből állnak. Így tehát két DNS-szál gerince között (amelyek váltakozó dezoxiribózból és foszforsavból állnak) minden különböző szálú nukleotidpár esetén mindig három gyűrű található (mivel a kétgyűrűs purin mindig csak egy bizonyos egygyűrűs gyűrűvel komplementer pirimidin). Ez lehetővé teszi, hogy a DNS-molekula szálai közötti szélesség végig azonos maradjon (körülbelül 2,3 nm).
A hélix egy fordulójában körülbelül 10 nukleotid található. Egy nukleotid hossza körülbelül 0,34 nm. A DNS-molekulák hossza általában óriási, meghaladja a nukleotidok millióit. Ezért annak érdekében, hogy tömörebben illeszkedjen a sejtmagba, a DNS különböző fokú "szupertekeredés"-en megy keresztül.
Amikor információt olvasunk a DNS-ből (vagyis az RNS-t szintetizáljuk rajta), ezt a folyamatot ún átírás) despiralizáció következik be (fordított spiralizációs folyamat), a két lánc egy speciális enzim hatására szétválik. A hidrogénkötések gyengék, ezért a láncok szétválása, majd térhálósodása alacsony energiafogyasztás mellett megy végbe. Az RNS-t a DNS-en szintetizálják a komplementaritás ugyanazon elve szerint. Csak az RNS-ben lévő timin helyett az uracil komplementer az adeninnel.
Genetikai kód A DNS-molekulákra írt hármasokból áll (három nukleotid szekvenciája), amelyek egy aminosavat (fehérje monomer) képviselnek. A DNS nagy része azonban nem kódol fehérjét. A molekula ezen részeinek jelentősége eltérő, sok tekintetben nem teljesen érthető.
A sejtosztódás előtt mindig megduplázódik a DNS mennyisége. Ezt a folyamatot ún replikáció... Természetében félig konzervatív: egy DNS-molekula láncai eltérnek egymástól, és mindegyiknek megvan a maga új kiegészítő lánca. Ennek eredményeként egy kétszálú DNS-molekulából két kettős szálú DNS-t kapunk, amelyek megegyeznek az elsővel.
A DNS-ben a polinukleotid láncok többirányúak, vagyis az egyik láncnak 5 "vége van (a foszforsavmaradék a dezoxiribóz ötödik szénatomjához kapcsolódik), a másiknak pedig a 3" vége van (foszforsavtól mentes). .
A replikáció és a transzkripció során a szintézis mindig az 5"végtől a 3-as felé haladó irányban halad, mivel az új nukleotidok csak a szabad 3"-os szénatomhoz tudnak kapcsolódni.
A DNS, mint örökletes információért felelős anyag szerkezete és szerepe a XX. század 40-50-es éveiben tisztázódott. 1953-ban D. Watson és F. Crick azonosította a DNS kétszálú szerkezetét. Korábban E. Chargaff rájött, hogy a DNS-ben a timin mennyisége mindig az adeninnek, a guanin mennyisége pedig a citozinnak felel meg.
Hasonló cikkek
-
Kínai nyelv - szövegek kezdőknek
Téma: Kína Számos olyan tulajdonsággal rendelkezik, amelyek kiemelik a többi ország közül. Először is ez a legnépesebb ország, és emiatt nagy számban vannak kínaiak, akik más országokba költöznek és ott telepednek le...
-
német záradékok
Németül kétféleképpen kell feltüntetni. Hasonlítsa össze: Er geht nach Deutschland, um Deutsch zu lernen. - Németországba megy németül tanulni. Ich schenke ihm ein deutsches Buch, damit er deutsche Literatur im Original liest. - Adok ...
-
Alárendelt szakszervezetek németül Damit és um zu németül
Sok német nyelvű diáknak sikerült már tollat, ceruzát, billentyűzetet, iPhone-t és laptopot összetörnie. Egyesek számára hihetetlenül bonyolultnak tűnik, másoknak éppen ellenkezőleg, egészen egyszerűnek. Egy dolog teljesen világos – a német nyelvben vannak dolgok...
-
Hogyan kezdjük el az orosz oktatást
Több hónapja tanulsz oroszul, de még mindig nem tudsz egy szót sem szólni, és a filmnézésről sem tudsz beszélni? Valószínűleg egyszerűen rosszul tanulsz oroszul. Beszéljünk az egyszerű, de hatékony szabályokról, amelyek...
-
A jármű pillanatnyi sebessége
A test gördítése ferde síkban (2. ábra); Rizs. 2. A test gördítése ferde síkban () Szabadesés (3. ábra). Mindez a három mozgástípus nem egységes, vagyis a sebesség változik bennük. Ebben a leckében mi...
-
(lehet valami univerzális képlet?
Az egységvektor olyan vektor, amelynek abszolút értéke (modulusa) egyenlő eggyel. Az egységvektor jelölésére az e alsó indexet használjuk. Tehát ha egy a vektor adott, akkor egységvektora az a e vektor lesz.